

导读:
结直肠癌是全球高发的恶性肿瘤,其恶性进展与异常的蛋白质稳态密切相关。近年来,靶向蛋白质合成已成为抗癌研究的热点,但如何精准打击癌细胞特有的合成依赖,同时避免伤及正常细胞,仍是巨大挑战。

本项研究,从一个传统中药材相关的天然化合物——去氧紫草素入手,系统揭示了其强效抑制结直肠癌的全新机制。DSHK能够作为一种“分子楔子”,精准插入并破坏癌细胞中两个关键蛋白HSPA8和GEMIN5所形成的功能复合体。这一破坏产生了灾难性的连锁反应:同时攻击“核糖体生成”和“翻译起始”!这双重机制,被定义为“剪接-翻译耦合”干预策略。
该研究不仅首次明确了DSHK的直接作用靶点,也揭示了一个全新的抗癌作用轴,为克服结直肠癌的异质性和耐药性提供了创新思路。
文章索引:
标题:Disruption of HSPA8-GEMIN5 interaction suppresses colorectal cancer by impaired splicing-translation coupling-mediated proteostasis imbalance
发表期刊:《Journal of Experimental & Clinical Cancer Research》.
发表时间:2026.01
作者团队:北京中医药大学 胡仲冬教授团队
IF:12.8
DOI:10.1186/s13046-026-03645-2.
研究概览
翻译组学技术应用:
l Polysome Profiling
用于分析DSHK处理后核糖体的组装状态。这直接证明了核糖体生物合成/组装过程的缺陷,是导致翻译能力下降的重要原因之一。
研究结果
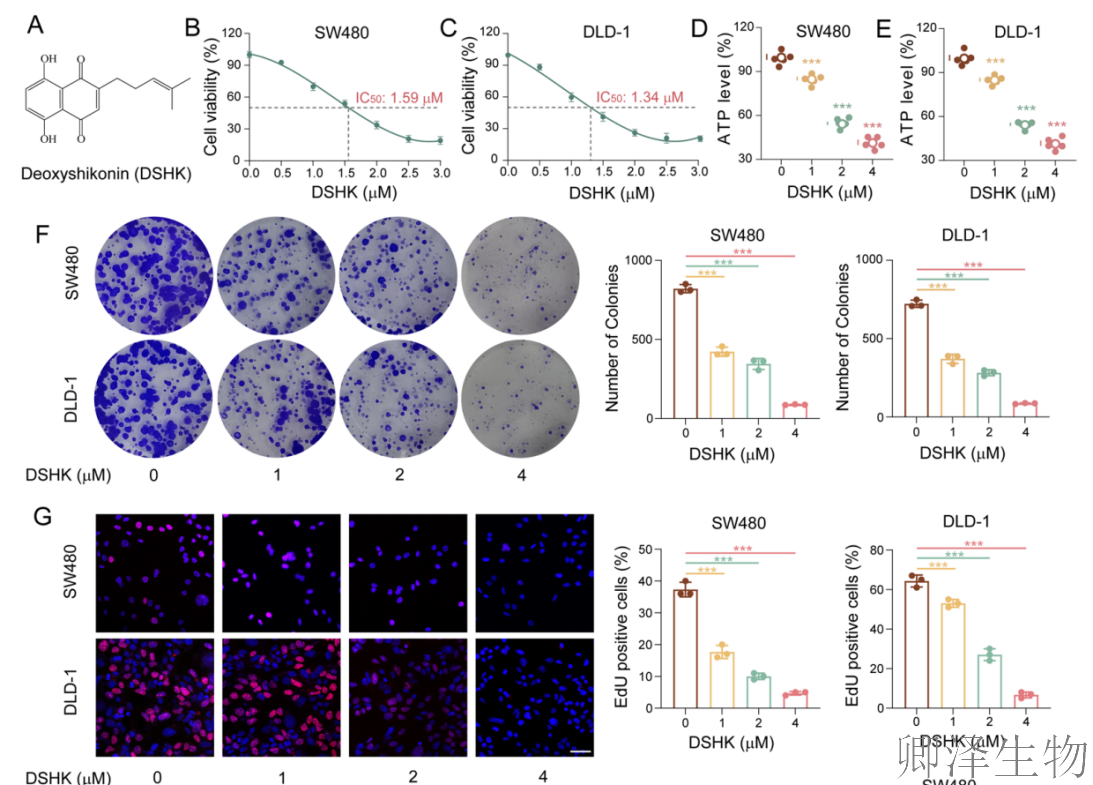
(1)DSHK在临床前结直肠癌模型中展现出显著的抗肿瘤活性
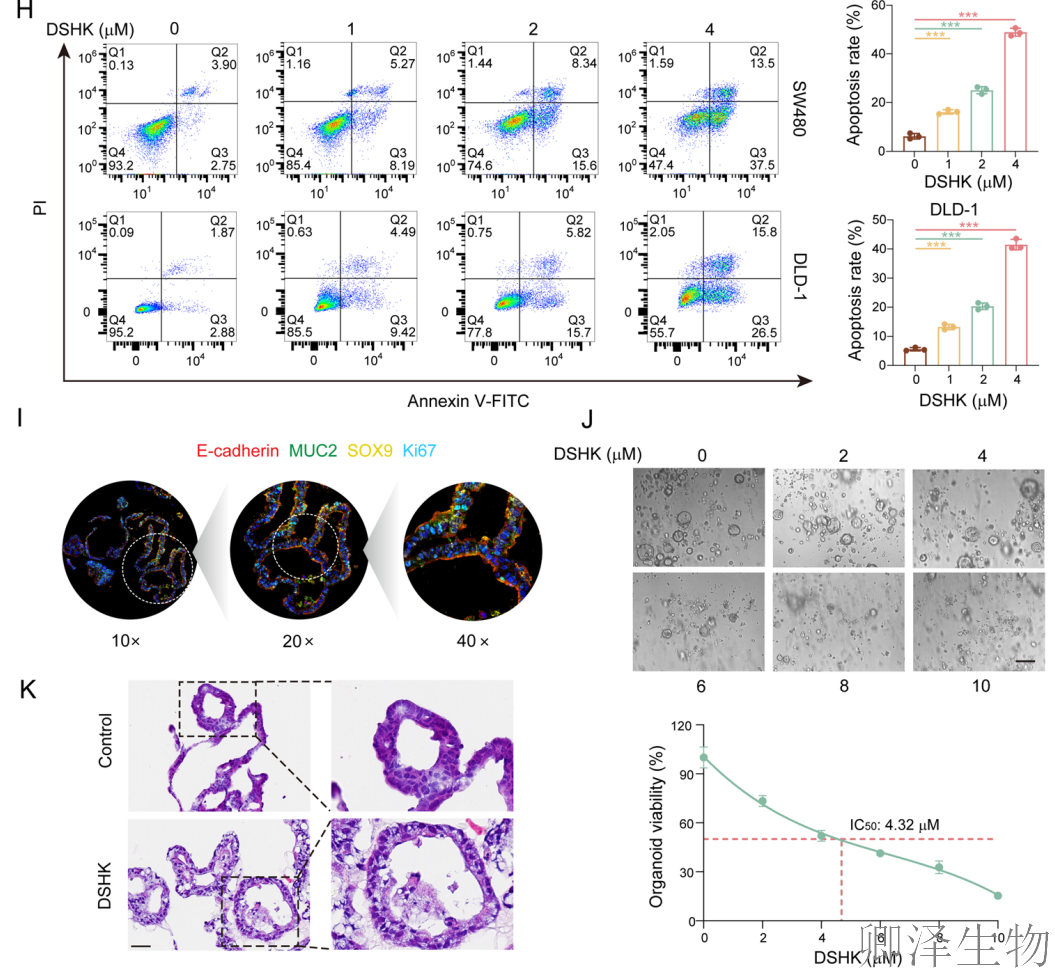
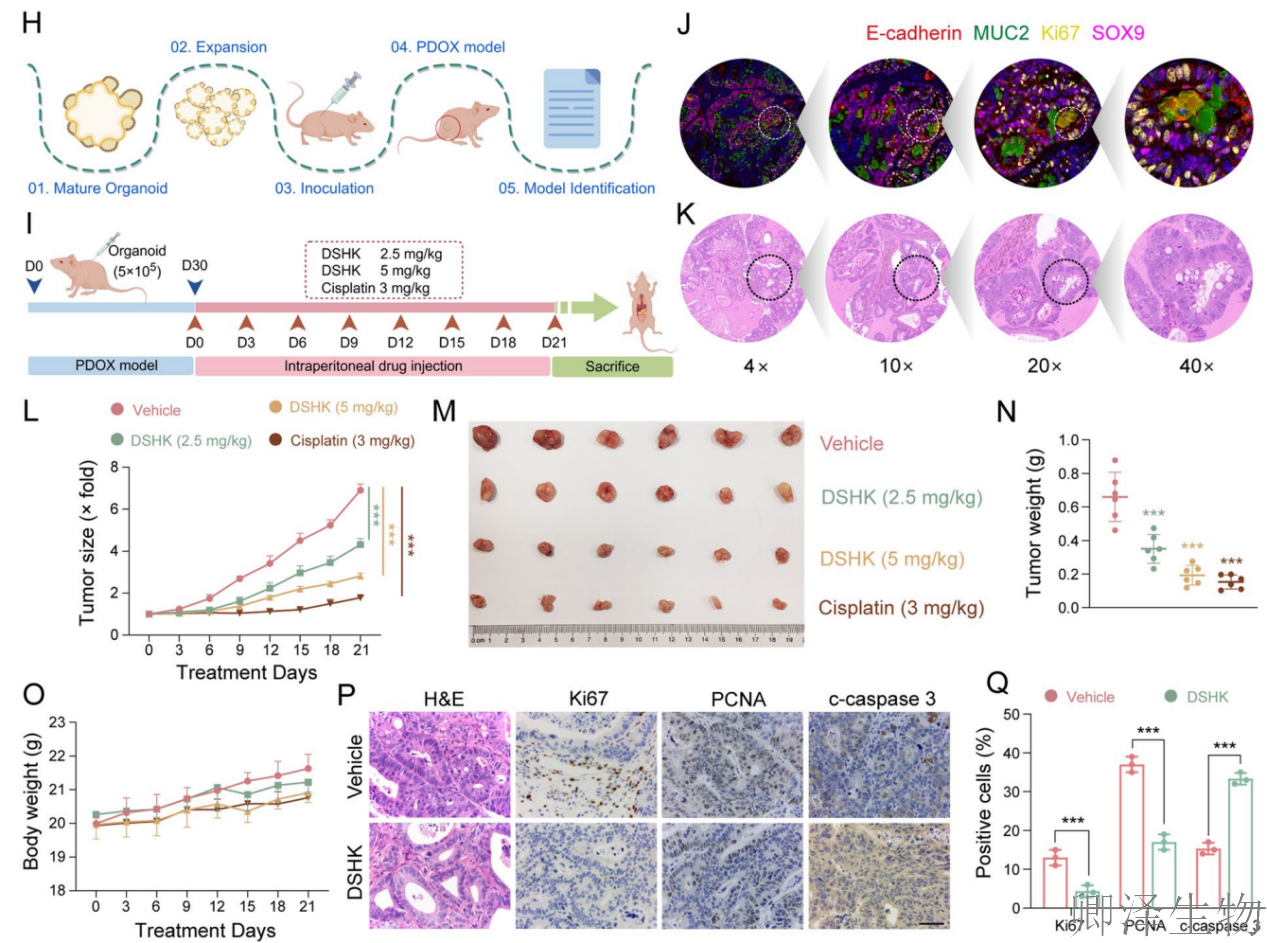
首先,作者通过系列实验建立了坚实的表型基础:天然化合物DSHK在多种临床前CRC模型中(包括细胞系和患者来源类器官)均展现出显著的抗肿瘤活性,其机制涉及抑制细胞能量代谢、阻断增殖并诱导凋亡。更重要的是,其对CRC细胞的毒性显著高于正常结肠细胞,揭示了其良好的肿瘤选择性(图1)。
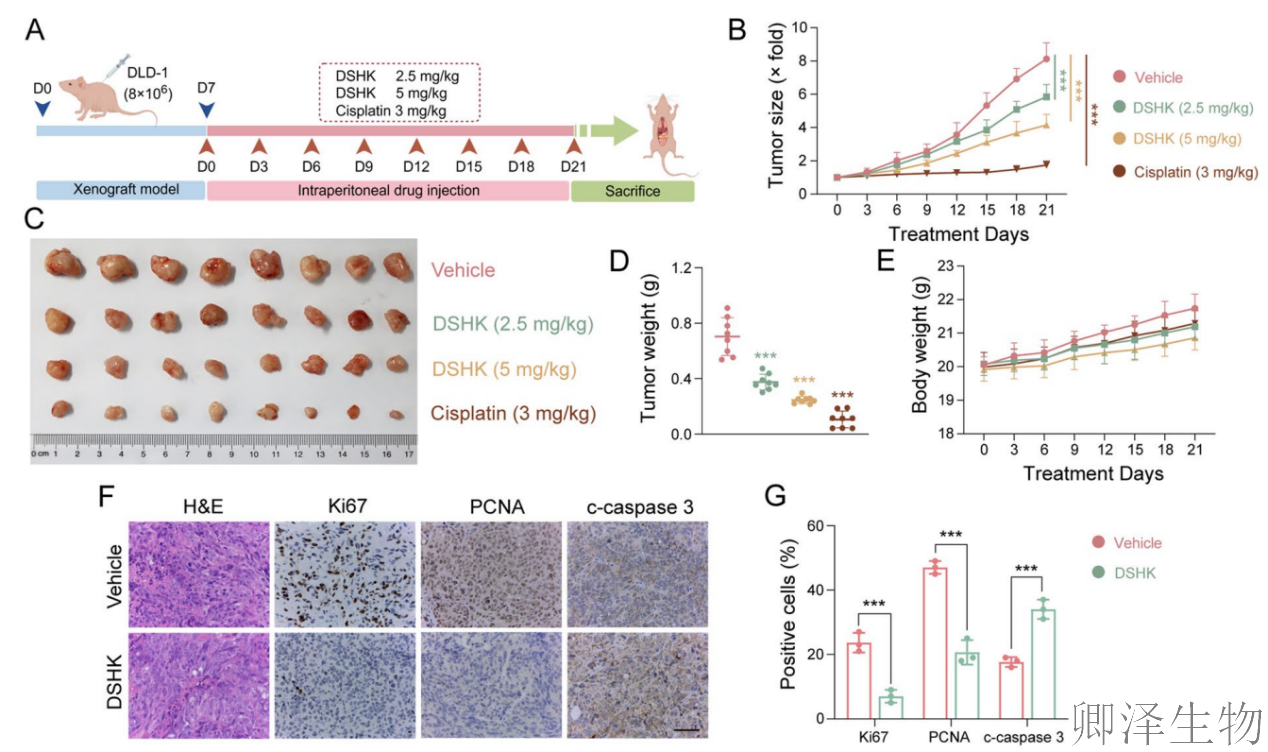
另外,在动物整体水平上成功验证了DSHK的治疗潜力(图2)。它不仅在不同类型的移植瘤模型(CDX和PDOX)中均表现出强效且剂量依赖性的抗肿瘤活性,能深度抑制肿瘤生长、改变其增殖与凋亡状态,更在有效剂量下展现出了良好的安全性。
(2)HSPA8是DSHK抗结直肠癌的直接靶点
接着,作者完成了一个教科书般的靶点鉴定工作。
首先,采用化学蛋白质组学策略,筛选到热休克蛋白家族(HSP)可能是DSHK的作用目标(图3A-C)。热休克蛋白重折叠实验,发现DSHK能剂量和时间依赖性地抑制HSP的修复功能,确证其靶向HSP通路。关键的细胞热转移实验显示DSHK特异性地与HSPA8在细胞内结合(图3D-G)。
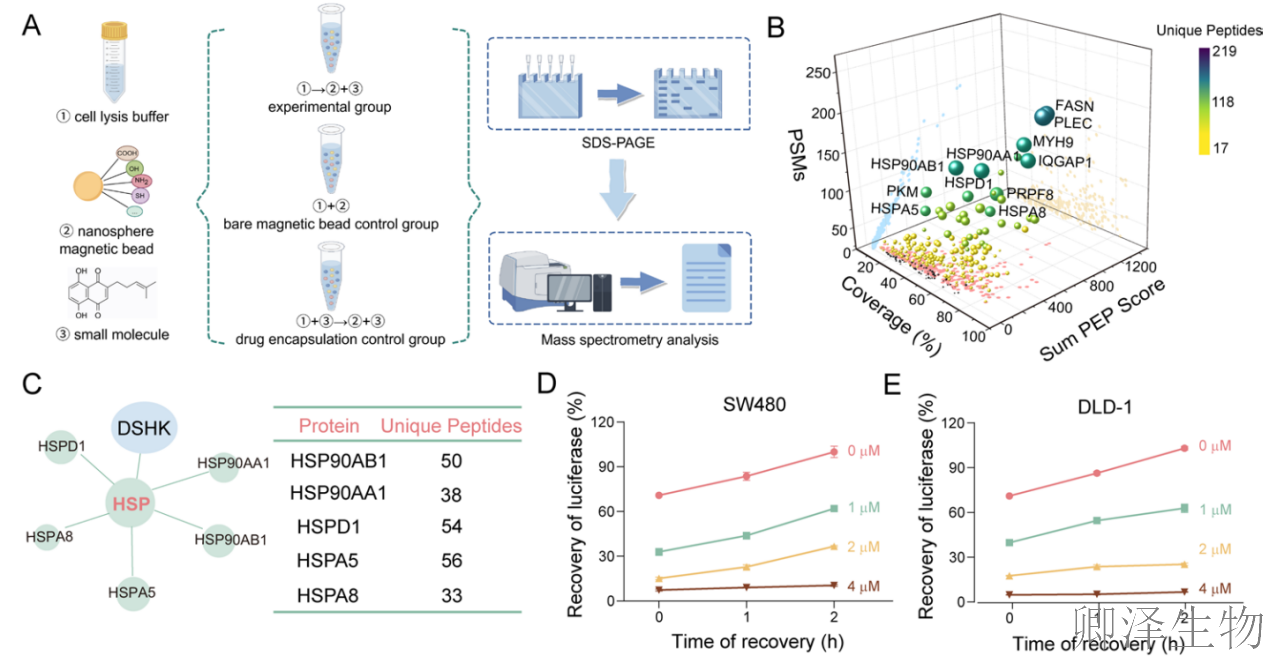
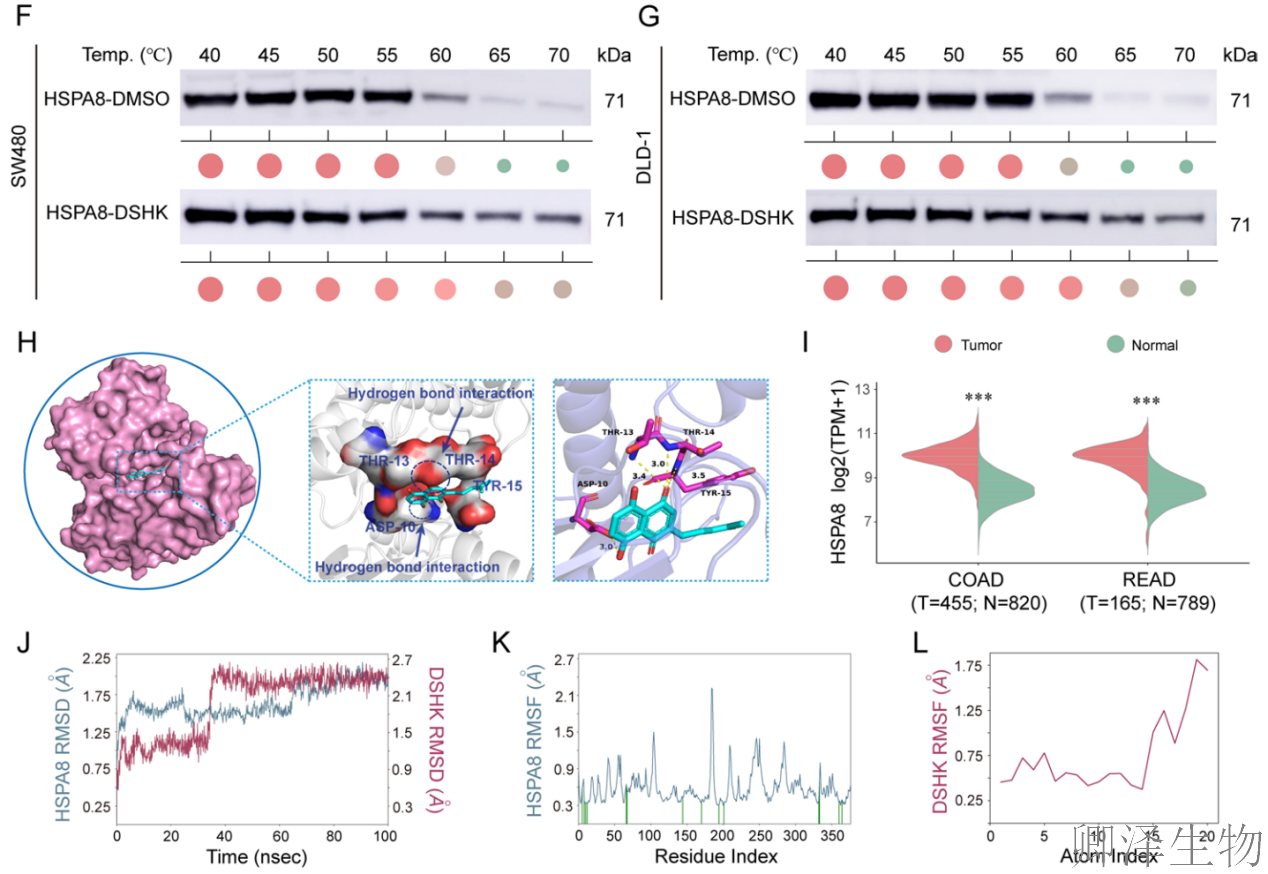
分子对接模拟和生物信息学分析再次强化了靶点依据(图3H-I)。同时,分子动力学模拟证明DSHK-HSPA8复合物构象非常稳定。表面等离子共振实验证实了可测量的、特异性的直接结合(图3J-M)。
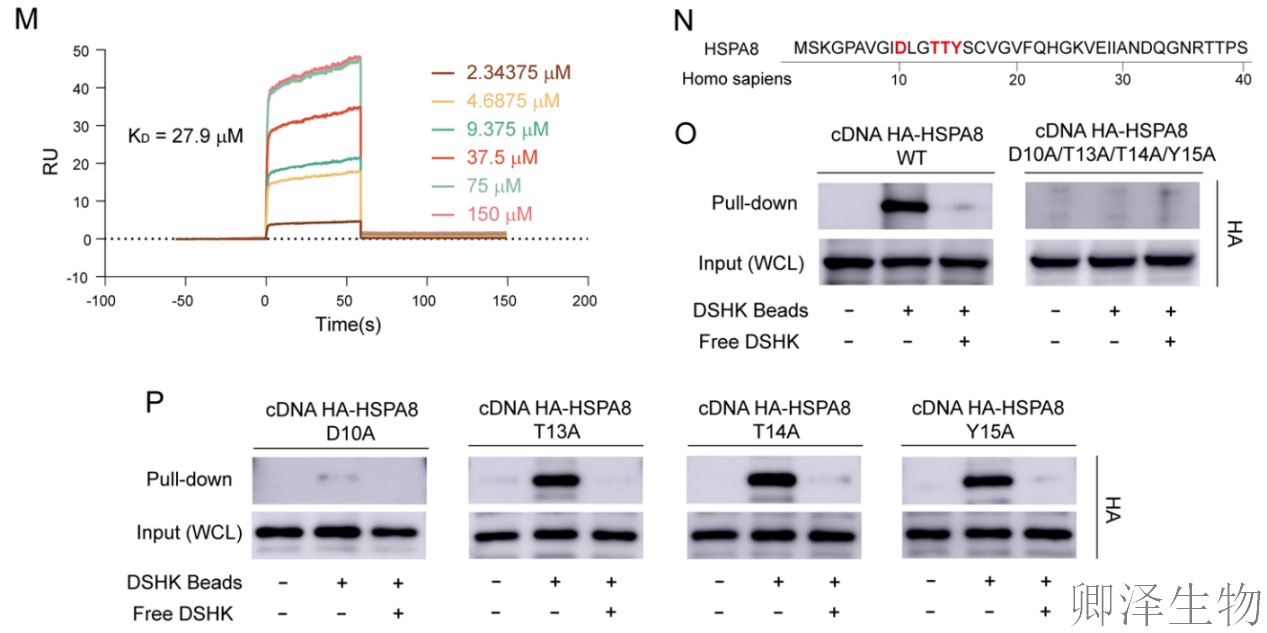
最关键的证据来自突变体结合实验(图3N-P)。将预测的结合位点残基突变为丙氨酸。结果发现,将四个残基同时突变,或仅单独突变核心位点ASP10,都会完全废除DSHK与HSPA8的结合。上述证据将HSPA8确立为DSHK抗CRC作用的直接分子靶标。
(3)DSHK的抗CRC活性取决于HSPA8
那DSHK的抗癌效果,是否必须通过结合并影响HSPA8才能实现?
在CRC细胞系和患者来源类器官中,HSPA8缺失本身就能显著抑制CRC细胞的增殖(图4A-E)。更关键的发现是,在HSPA8敲低的细胞和类器官中,DSHK抑制细胞活力、降低ATP水平以及阻断集落形成的能力被大幅削弱(图4F-J)。
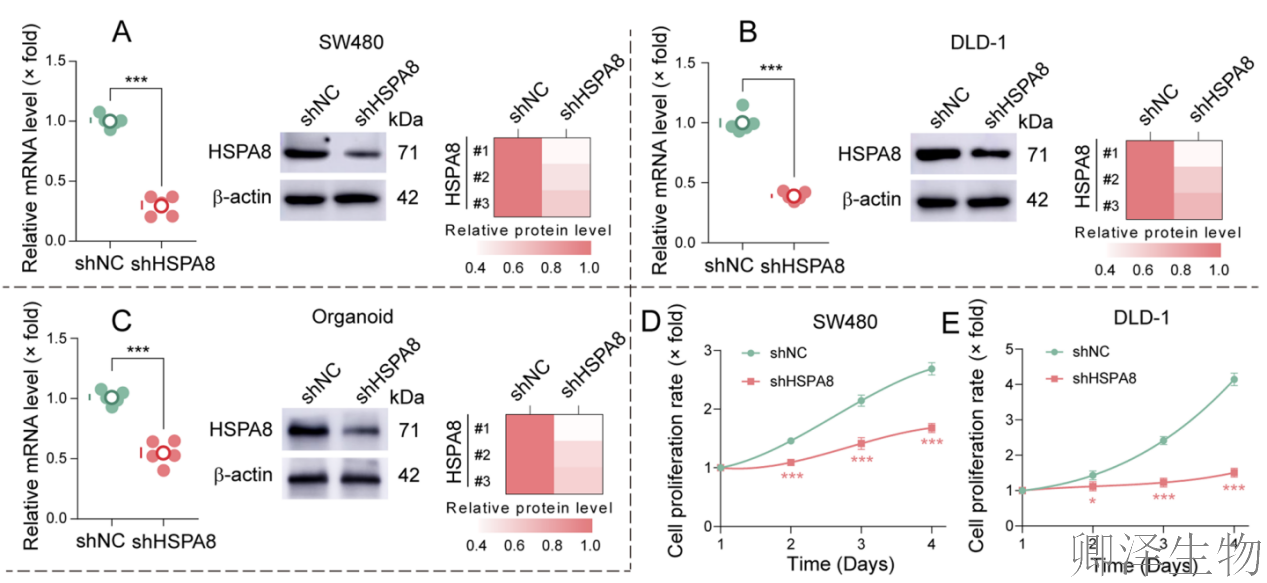
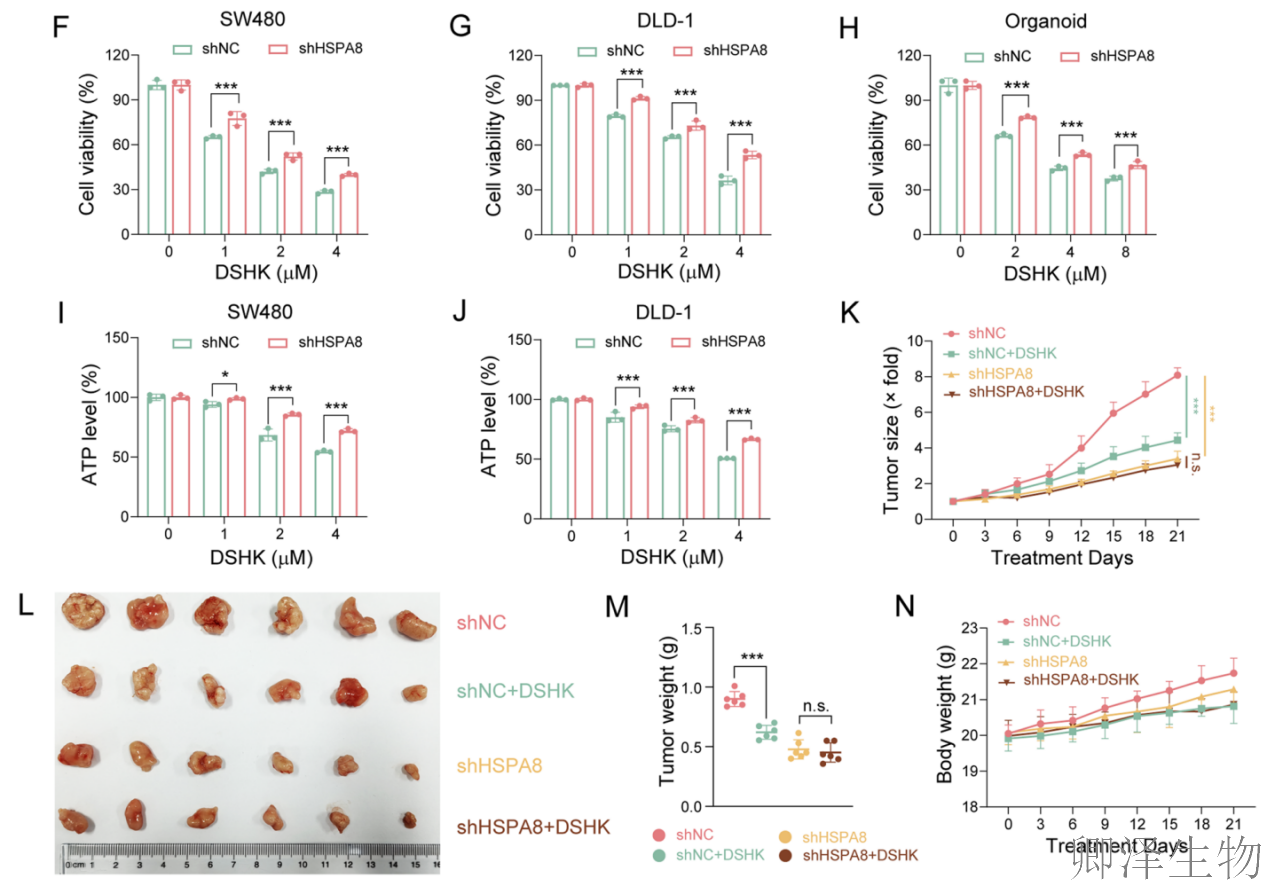
最有力的证据来自体内实验(图4K-N)。将稳定敲低HSPA8的CRC细胞移植到小鼠体内形成肿瘤。在HSPA8敲低组,DSHK治疗完全失去了抑制肿瘤生长的能力。这与对照组(野生型细胞)中DSHK显著的抑瘤效果形成鲜明对比。这无可辩驳地证明,在复杂的生物体内,DSHK的抗肿瘤疗效也完全依赖于其靶蛋白HSPA8的存在。
(4)GEMIN5是HSPA8的新型功能相互作用子
那HSPA8究竟通过和哪些下游蛋白“搭档”来驱动CRC进展?
为了全面绘制HSPA8在CRC细胞中的相互作用图谱,作者使用了SPIDER邻近标记技术(图5A-B)。质谱分析筛选出四个高置信度的HSPA8结合蛋白,包括GEMIN5。分子对接模拟预测HSPA8与GEMIN5之间存在较强的结合能和互补的相互作用界面(图5C-D, J)。更重要的是,TCGA数据分析显示,GEMIN5在CRC肿瘤组织中显著高表达,且高表达与患者不良预后相关。
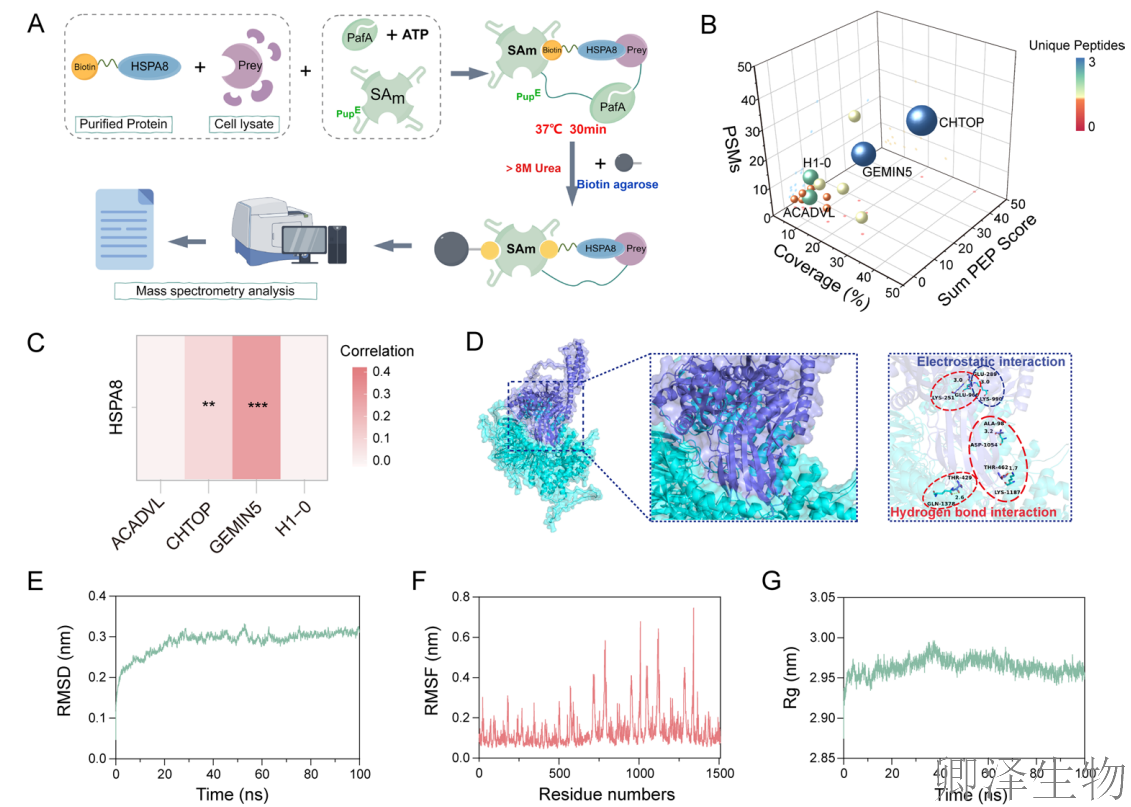
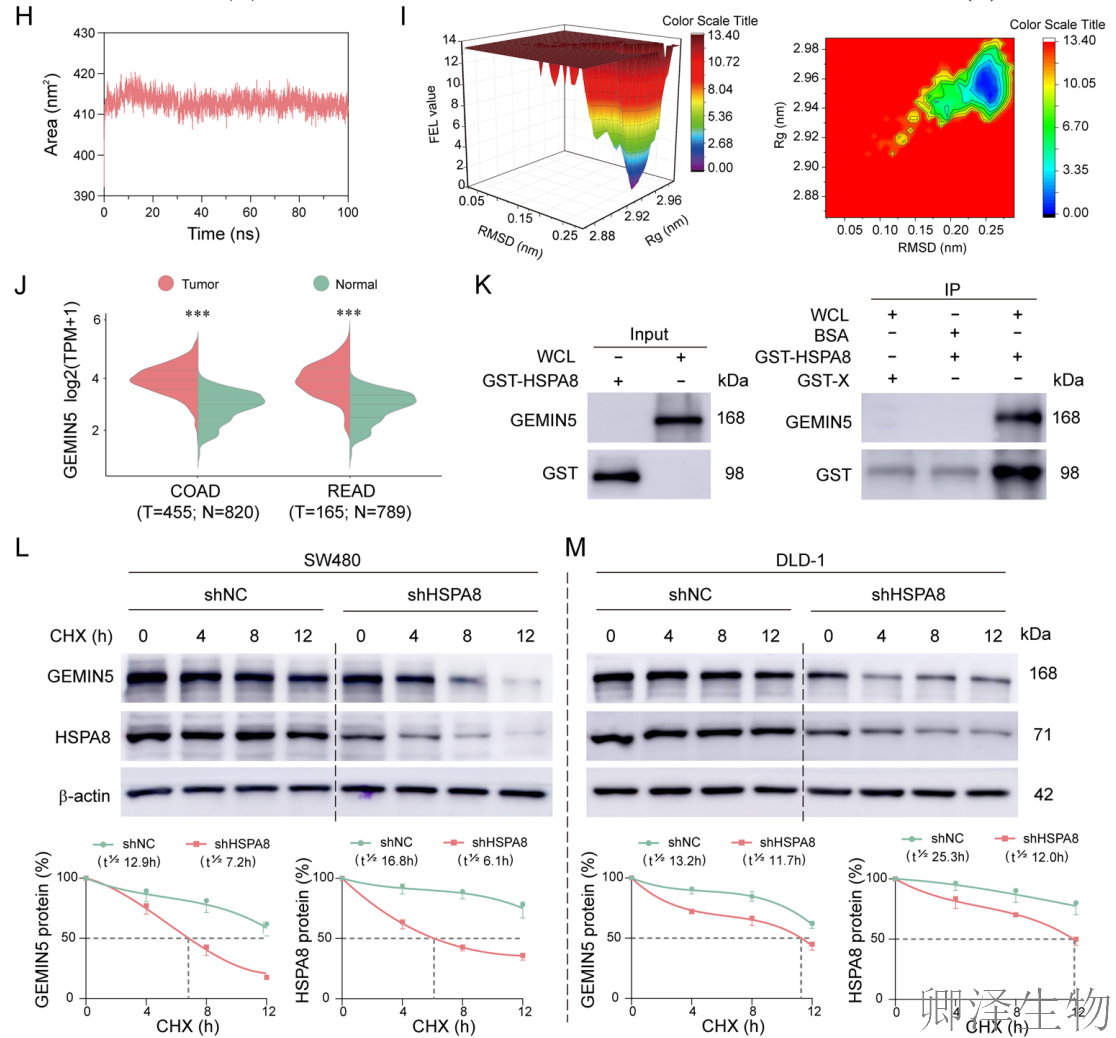 为了在原子层面评估互作的可靠性,作者进行了分子动力学模拟(图5E-I)。从计算物理学角度强有力地支持二者可形成稳定的复合物。GST pull-down实验直接证实(图5K),纯化的重组HSPA8蛋白能够特异性地“拉下”细胞裂解液中的GEMIN5,提供了二者直接物理相互作用的生化学证据。
为了在原子层面评估互作的可靠性,作者进行了分子动力学模拟(图5E-I)。从计算物理学角度强有力地支持二者可形成稳定的复合物。GST pull-down实验直接证实(图5K),纯化的重组HSPA8蛋白能够特异性地“拉下”细胞裂解液中的GEMIN5,提供了二者直接物理相互作用的生化学证据。
最关键的功能性证据来自蛋白稳定性实验(图5L-M)。当在CRC细胞中敲低HSPA8后,GEMIN5蛋白的半衰期显著缩短。这表明HSPA8作为分子伴侣,对维持其互作蛋白GEMIN5的稳定性至关重要。
(5)DSHK破坏了HSPA8-GEMIN5相互作用
DSHK通过结合HSPA8,是否会像“分子楔子”一样,破坏这个致癌复合体的完整性?
在免疫共沉淀实验中,在CRC细胞中,内源性的HSPA8能有效“拉下”GEMIN5,反之亦然,确证了二者在生理环境下形成稳定复合物。而且,用DSHK处理细胞后,HSPA8与GEMIN5的共沉淀效率显著降低。
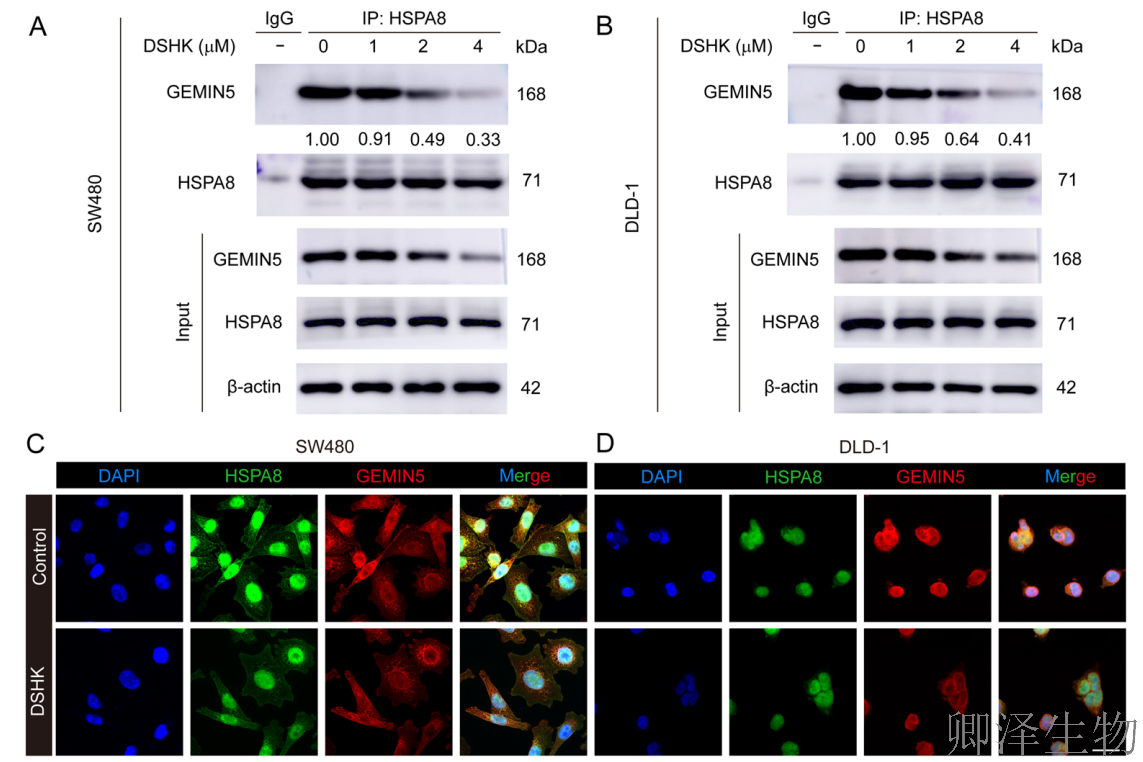
为了在更接近真实生理状态的空间环境中验证这一破坏,作者在三种模型中进行了免疫荧光共定位分析(图6C-L):在CRC细胞系(SW480/DLD-1)、患者来源类器官以及小鼠移植瘤组织中,均观察到HSPA8和GEMIN5在核周区域存在高度的空间共定位。当DSHK处理后,在上述所有模型中,HSPA8与GEMIN5荧光信号的空间重叠程度(Pearson相关系数)均显著下降。
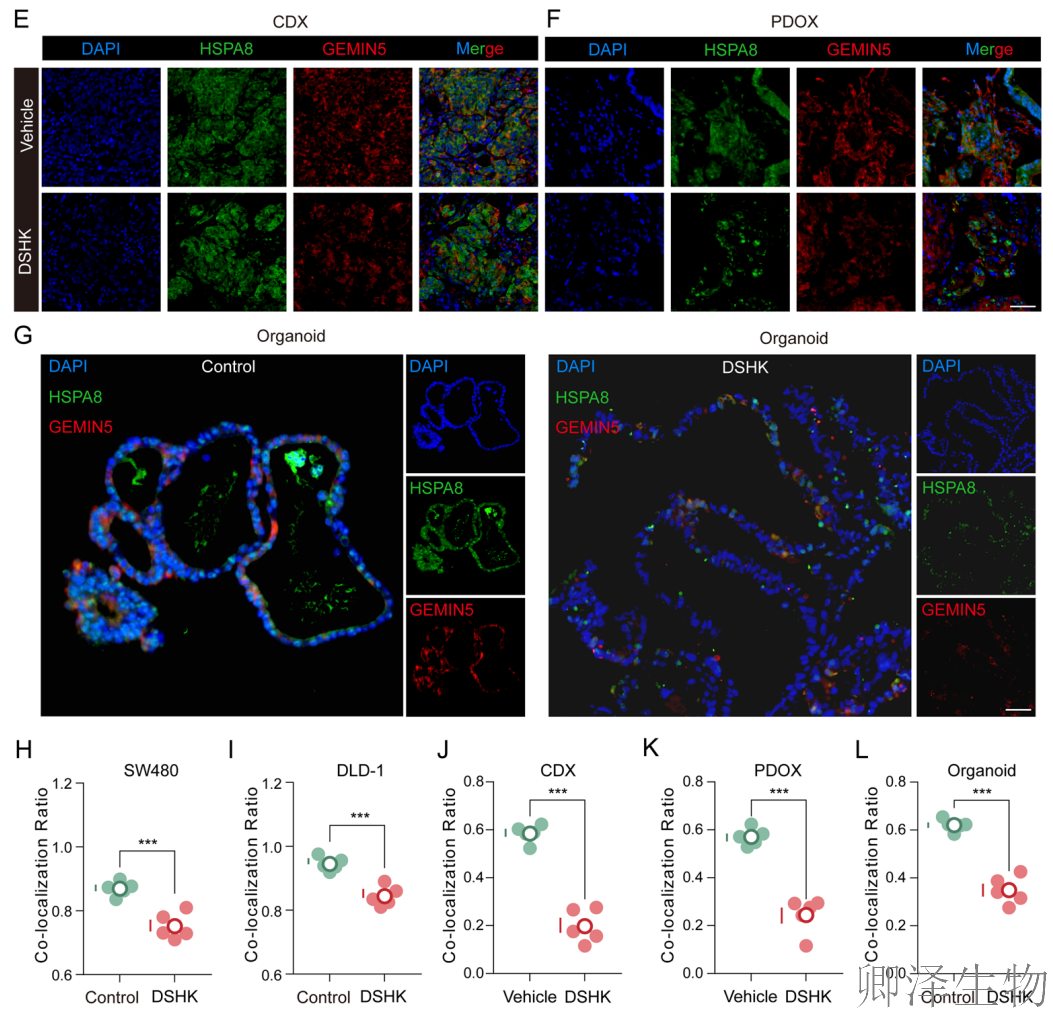
(6)HSPA8和GEMIN5表达及其相互作用在CRC中的临床相关性
随后,作者将基础研究发现“翻译”到了临床语境。它不仅在大量患者样本中证实了HSPA8和GEMIN5是CRC中协同高表达、空间共定位的致癌因子,更通过超分辨成像提供了二者在肿瘤中形成纳米尺度紧密复合物的直观证据。

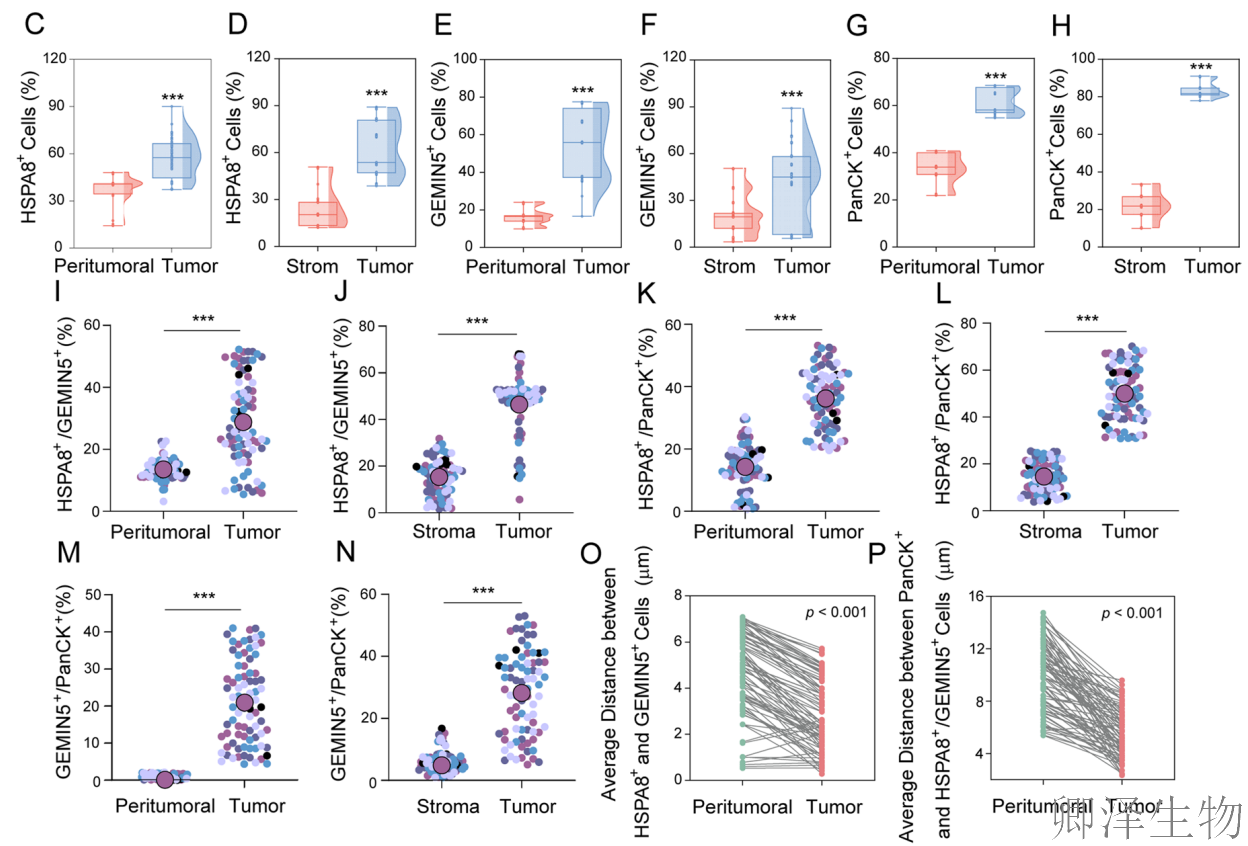
综合多个维度,强有力地证实了HSPA8-GEMIN5轴在CRC发生发展中的关键作用,并为针对该轴的靶向治疗提供了直接的临床合理性。
(7)DSHK通过GEMIN5功能损伤诱导RP编码基因的AS
GEMIN5是RNA剪接的关键调控因子。那DSHK导致的GEMIN5功能损伤,如何具体影响基因表达,并最终抑制蛋白质合成?
WB分析显示,DSHK处理不仅降低了GEMIN5自身的蛋白水平,也同步下调了其关键互作剪接因子U2AF2和ASF的表达(图8A-D)。对DSHK处理或HSPA8敲低细胞的RNA-seq数据进行全转录组可变剪接分析(图8E-G),发现两者均能显著增加基因组范围内的异常剪接事件总数。通过rMATS算法定量发现,所有七种主要类型的可变剪接事件频率均显著升高。
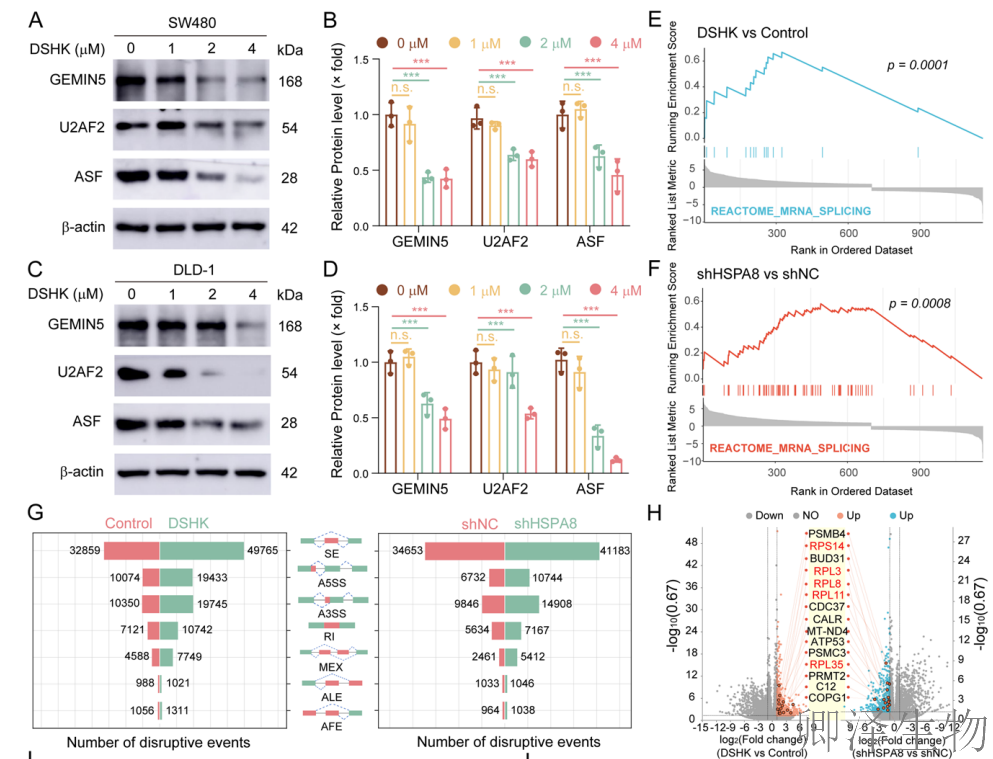
在众多发生异常剪接的基因中,作者通过差异分析筛选出15个受影响最显著的基因(图8H-J)。其中,5个是核糖体蛋白编码基因(RPL11, RPS14, RPL3, RPL35, RPL8)。这些基因的剪接错误主要集中在A3SS和A5SS类型。
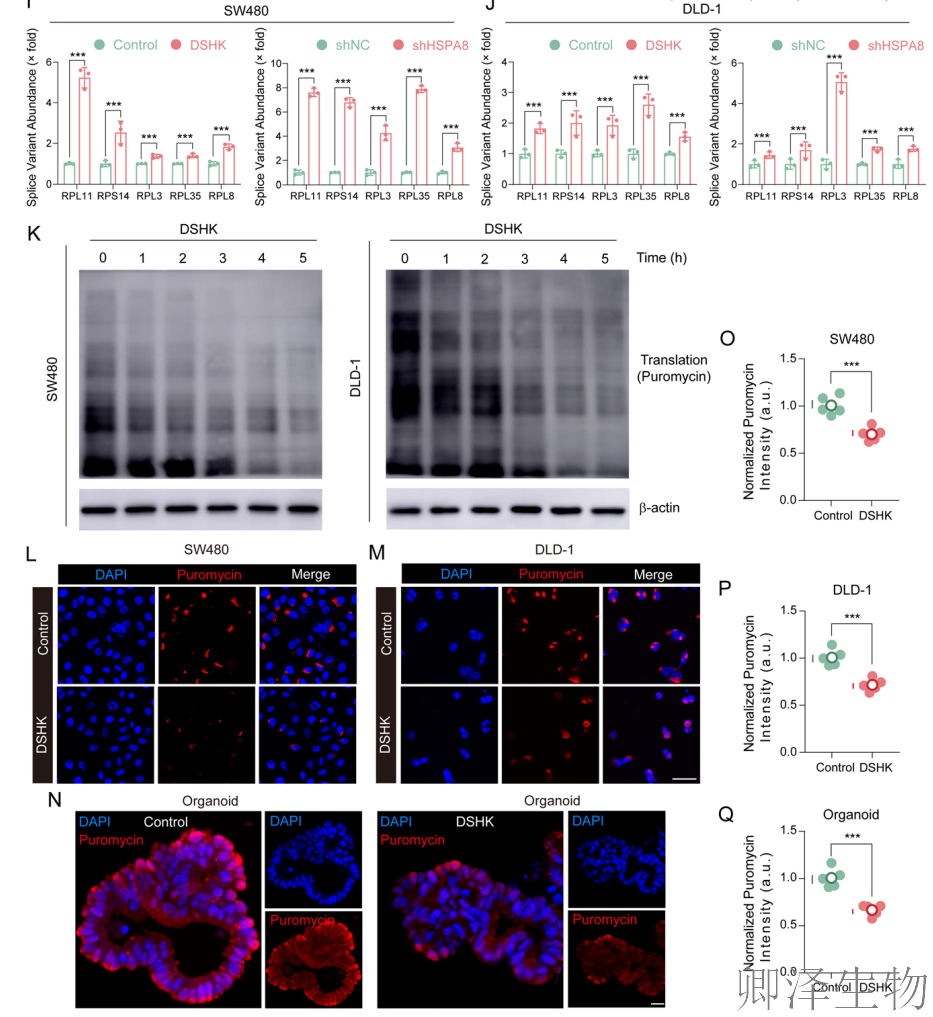
基于“有缺陷的核糖体蛋白将导致有缺陷的核糖体,进而损害翻译”的合理假设,作者直接检测了蛋白质合成速率(图8K-Q)。嘌呤霉素新生链掺入实验显示,DSHK处理能时间依赖性地强烈抑制SW480和DLD-1细胞以及患者来源类器官中的全局新生蛋白质合成。这从功能上确认了上游的剪接紊乱最终导致了蛋白质“生产线”的停工。
(8)DSHK阻碍功能性核糖体生物发生
透射电子显微镜捕捉到DSHK处理后导致“核糖体丢失症”(图9A-C)。这为“核糖体生物合成受损”提供了最直观、最震撼的形态学证据。
为了量化核糖体的状态,作者进行了Polysome profiling分析(图9D-E)。结果显示,DSHK处理的细胞中,80S核糖体单体峰显著减弱。同时,其前体40S和60S亚基的比例也发生失衡。这表明,不仅成熟核糖体总量减少,其组装过程也出现了故障。
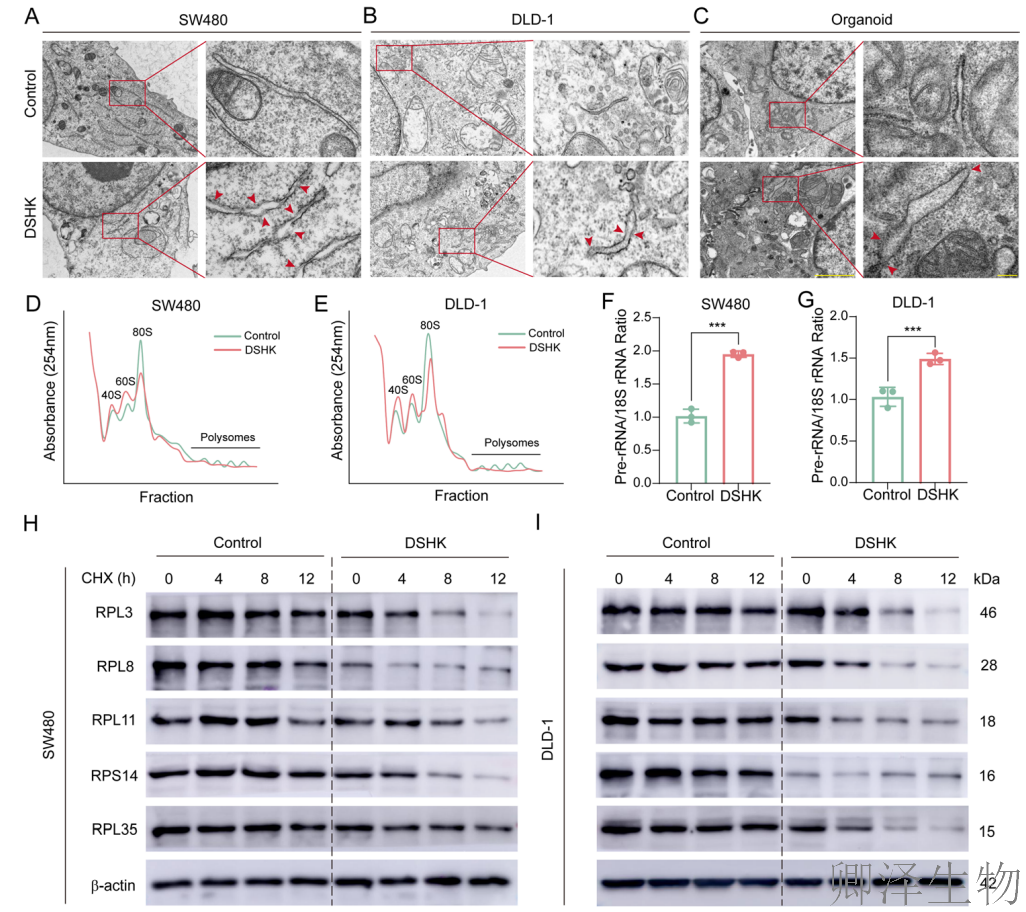
核糖体的生物合成需要rRNA前体的正确加工和成熟。DSHK处理后,前体rRNA与成熟18S rRNA的比值显著升高(图9F-G)。这说明核糖体RNA的加工成熟过程被阻断。
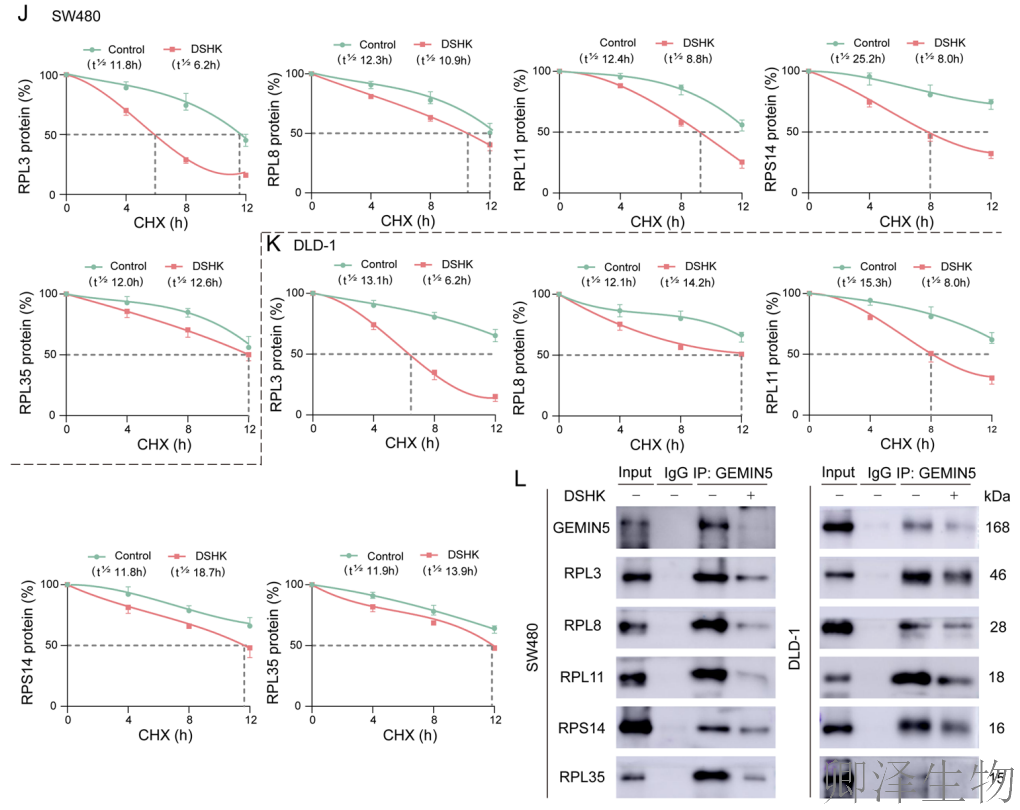
并且,蛋白稳定性实验(CHX追踪)证实,那些发生异常剪接的RP(如RPL3, RPL8等),其蛋白半衰期在DSHK处理后显著缩短(图9H-K)。Co-IP实验显示(图9L),DSHK处理显著削弱了GEMIN5与多个关键RP之间的相互作用。
综上,DSHK通过破坏HSPA8-GEMIN5轴,导致GEMIN5功能失常,进而引发一系列核糖体生物合成的灾难性后果——包括rRNA加工受阻、RP稳定性下降、RP与组装因子互作破坏,最终表现为核糖体亚基失衡、成熟核糖体锐减,并在电镜下呈现为广泛的“去核糖体化”。
(9)HSPA8介导DSHK诱导的翻译起始功能障碍
最后,作者阐明了DSHK抑制蛋白质合成的第二条并行通路:DSHK靶向结合HSPA8 → 破坏HSPA8与翻译起始因子(eIF4A/4G/4E)之间的相互作用 → 导致eIF4F翻译起始核心复合物组装失败与解离 → 触发翻译起始抑制信号(p-eIF2α升高) → 最终阻碍翻译起始。
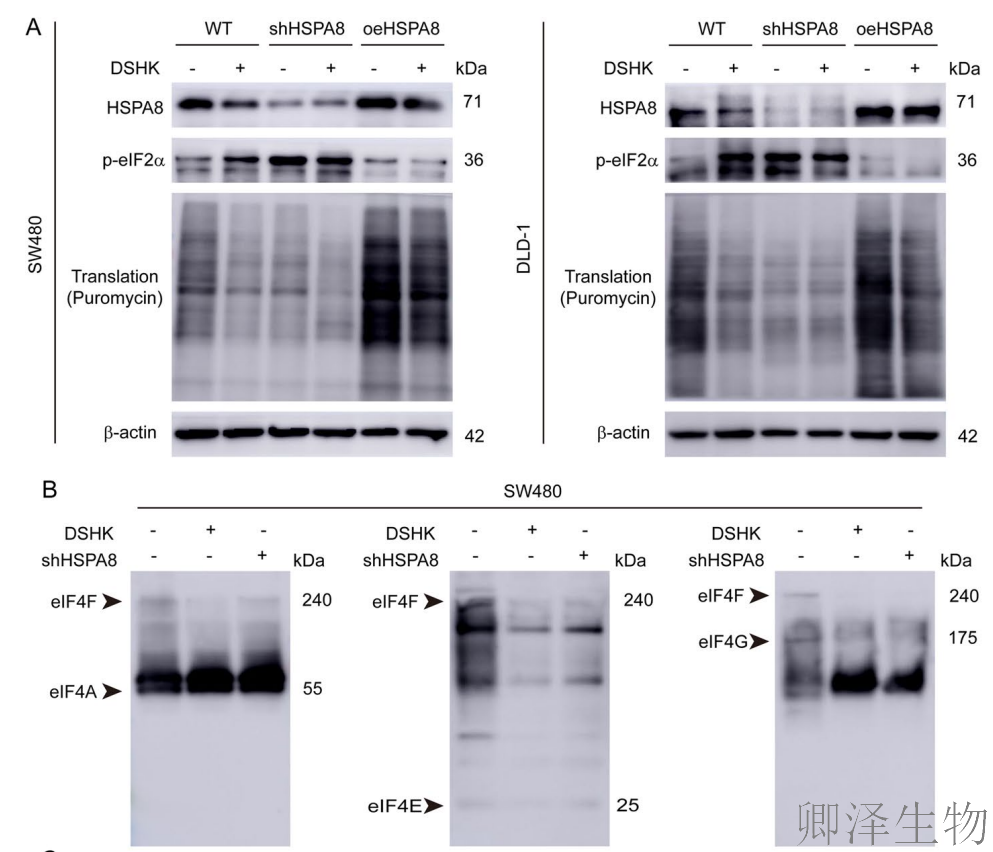
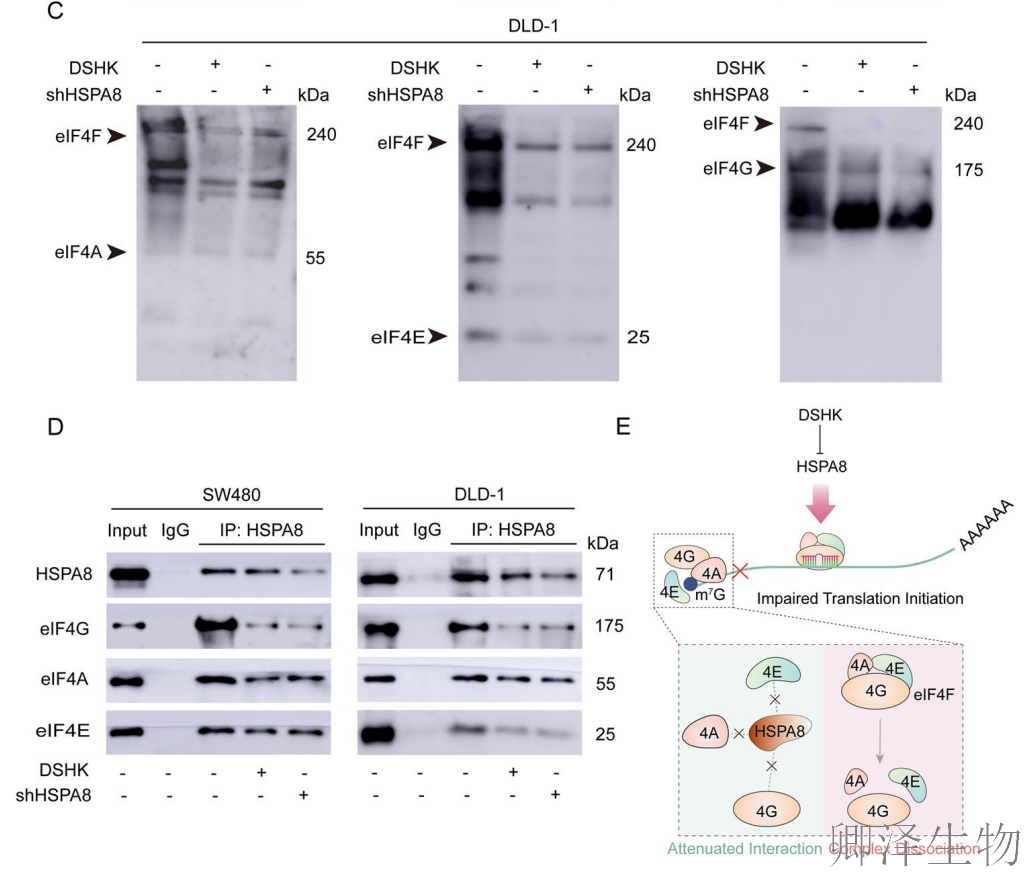
这项工作与上述揭示的“破坏核糖体生物合成”通路相辅相成,共同构成了DSHK通过靶向HSPA8-GEMIN5枢纽,对CRC细胞的蛋白质合成实施“上游起始抑制”与“下游机器破坏”的双重打击策略,完整地阐释了其强效抗癌作用的分子基础。
结论
本研究发现了一条全新的CRC治疗轴心:DSHK → 结合并破坏HSPA8-GEMIN5复合体 → 一方面通过GEMIN5损害剪接保真性,破坏核糖体生物合成(“毁坏生产机器”);另一方面通过HSPA8抑制翻译起始复合体组装(“关闭启动开关”)→ 全局蛋白质合成崩溃 → 抑制肿瘤生长。该研究将HSPA8-GEMIN5确立为介导“剪接-翻译耦合”的关键分子枢纽,为CRC提供了创新的“双通路干预”治疗策略。









