

你是否想过,癌细胞是如何在人体内“隐身”,逃避免疫系统追杀的?这背后,一个名为“无义介导的mRNA降解”(NMD)的细胞质量控制机制,可能扮演了关键角色。
此项研究揭开了肿瘤免疫逃逸的新面纱:一个名为LTO1/YAE1的蛋白质复合体,竟能通过调控NMD,来“下调”肿瘤细胞表面的“身份标签”——MHC-I分子,从而帮助癌细胞躲避免疫T细胞的识别。更令人兴奋的是,研究发现,一种常见的铁螯合剂,竟能巧妙“关闭”这个机制,让肿瘤重新“现身”,并显著提升免疫疗法的效果。这为攻克肿瘤免疫抵抗带来了全新希望。
文章索引:
标题:LTO1 and YAE1 regulate MHC-I expression via nonsense-mediated RNA decay in tumor cells
发表期刊:《Journal for ImmunoTherapy of Cancer》.
发表时间:2025.09
作者团队:细胞命运调控与疾病粤港澳高校联合实验室 王晓玲团队
IF:10.6
DOI:10.1136/jitc-2025-011893.
技术应用:
Polysome profiling
借助该技术直接评估了LTO1和YAE1敲除对细胞整体翻译状态的影响:翻译效率受损 + 核糖体生物合成缺陷
研究结果
LTO1/YAE1作为调节mRNA降解和翻译终止的NMD因子
作者在黑色素瘤细胞里安装了一套精巧的“双色荧光报告系统”:一个红色荧光蛋白基因后面跟着它的“终止密码子”,再后面是一个绿色荧光蛋白基因。正常情况下,细胞读到“终止密码子”就会停下,只产生红色荧光。但设计了一个小技巧,试图诱骗细胞自身的“编辑工具”把这个“终止符”改成一个“继续符”,如果成功,细胞就会产生一个红绿融合的荧光蛋白。
作者用这个系统对全基因组近2万个基因进行了大规模的“排查”,意外发现敲除LTO1或YAE1基因会导致绿色荧光显著增强,首次将它们锁定为潜在的调控分子(图1A-C)。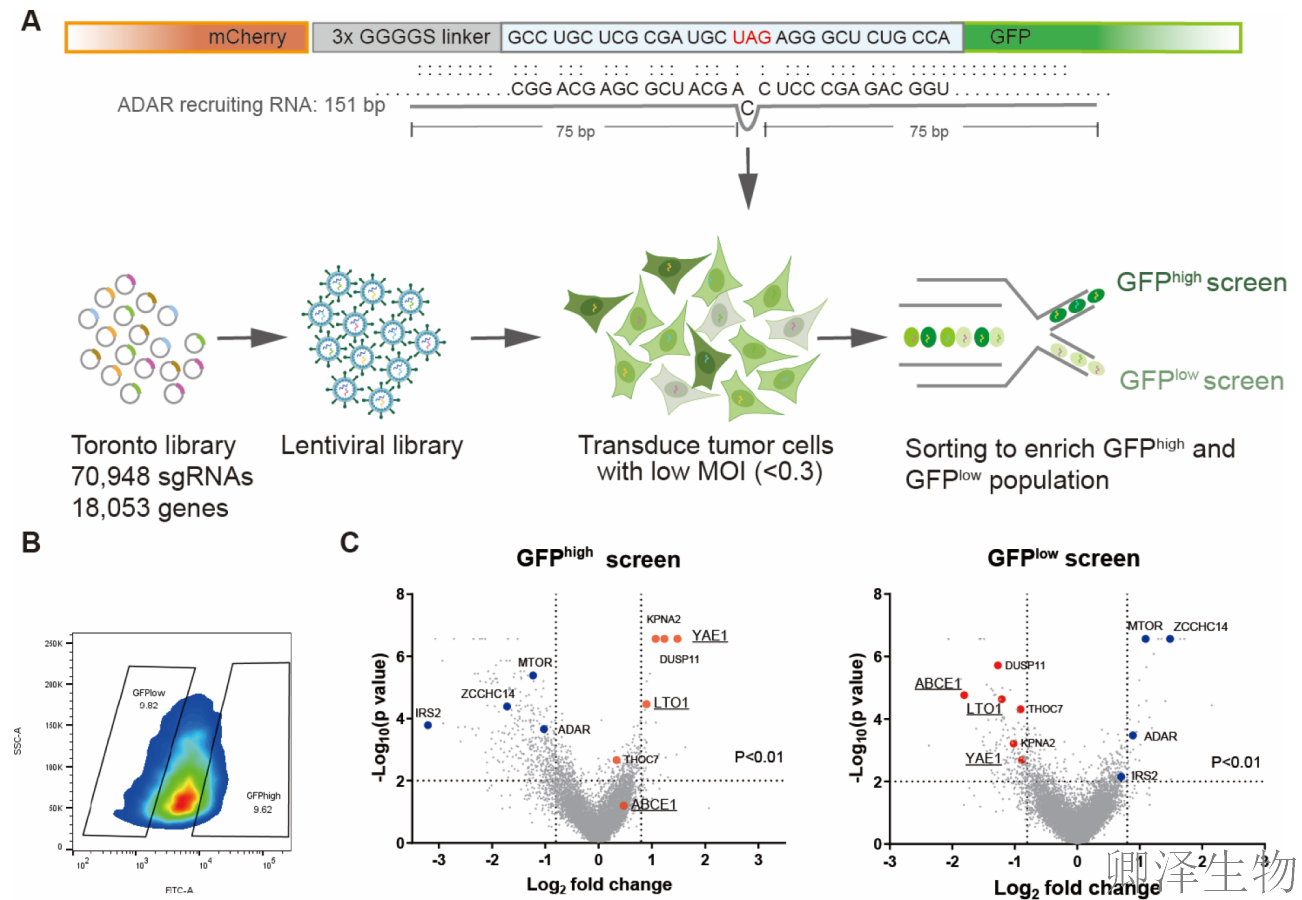
单独敲除LTO1或YAE1后,不仅绿色荧光蛋白(GFP)表达大幅上升,红色荧光蛋白(mCherry)表达也增加了,且两者来自同一条mRNA(图1D-G)。WB结果进一步显示,GFP蛋白的增加幅度远高于mCherry,并出现了非预期的较小GFP蛋白条带。这表明,在缺失LTO1/YAE1后,细胞的“翻译机器”可能异常地越过了正常的终止信号,并在下游非经典位置重新启动了翻译。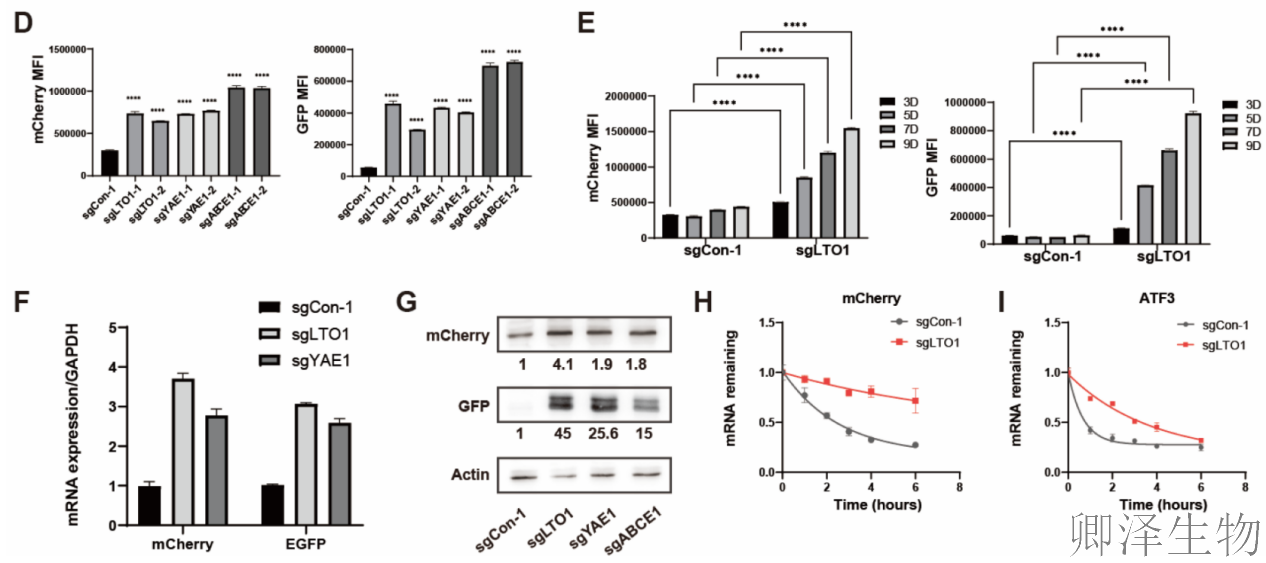
mRNA稳定性实验证明,在LTO1缺失的细胞中,报告基因mRNA和已知的NMD底物mRNA降解速度都显著变慢(图1H-I)。这证实了LTO1在“无义介导的mRNA降解”这一质量控制机制中扮演关键角色。
LTO1和YAE1是NMD和核糖体生物发生所必需的
随后,研究深入揭示LTO1和YAE1的分子功能。
转录组测序分析发现,敲除 LTO1、YAE1或ABCE1的细胞中,有大量基因表达发生共同变化,其中包含许多已知的NMD靶标基因(图2A-C)。这直接证明这三个基因协同作用,广泛参与调控细胞的“mRNA质量监控(NMD)”系统。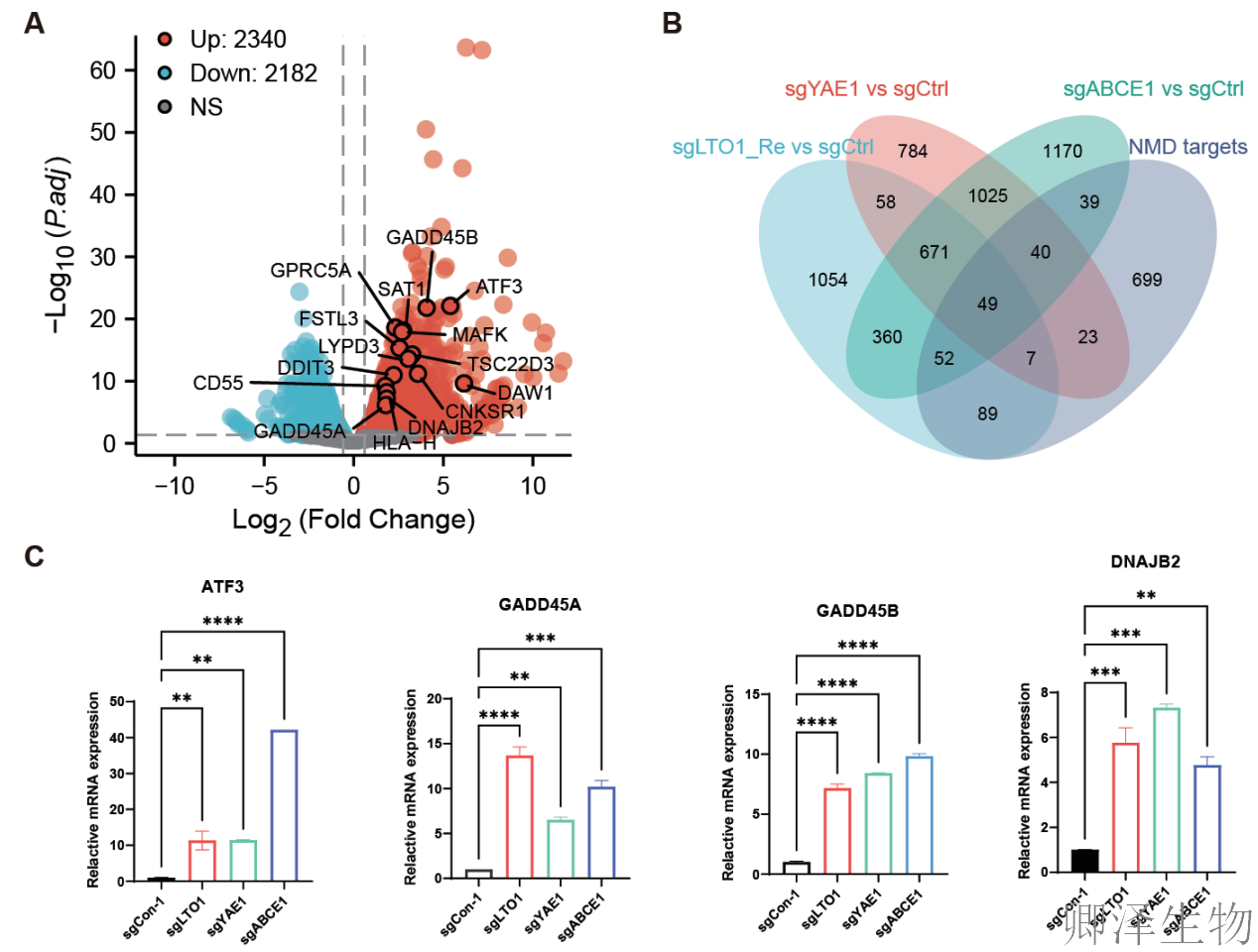
进一步的生物信息学分析和实验验证揭示(图2D-F),LTO1/YAE1的功能远不止于NMD。它们还深度参与“蛋白质合成工厂”——核糖体的组装。当它们缺失时,负责构建核糖体60S大亚基的关键蛋白(如RPL7, RPL14)产量显著下降,暗示核糖体“生产线”出现了问题。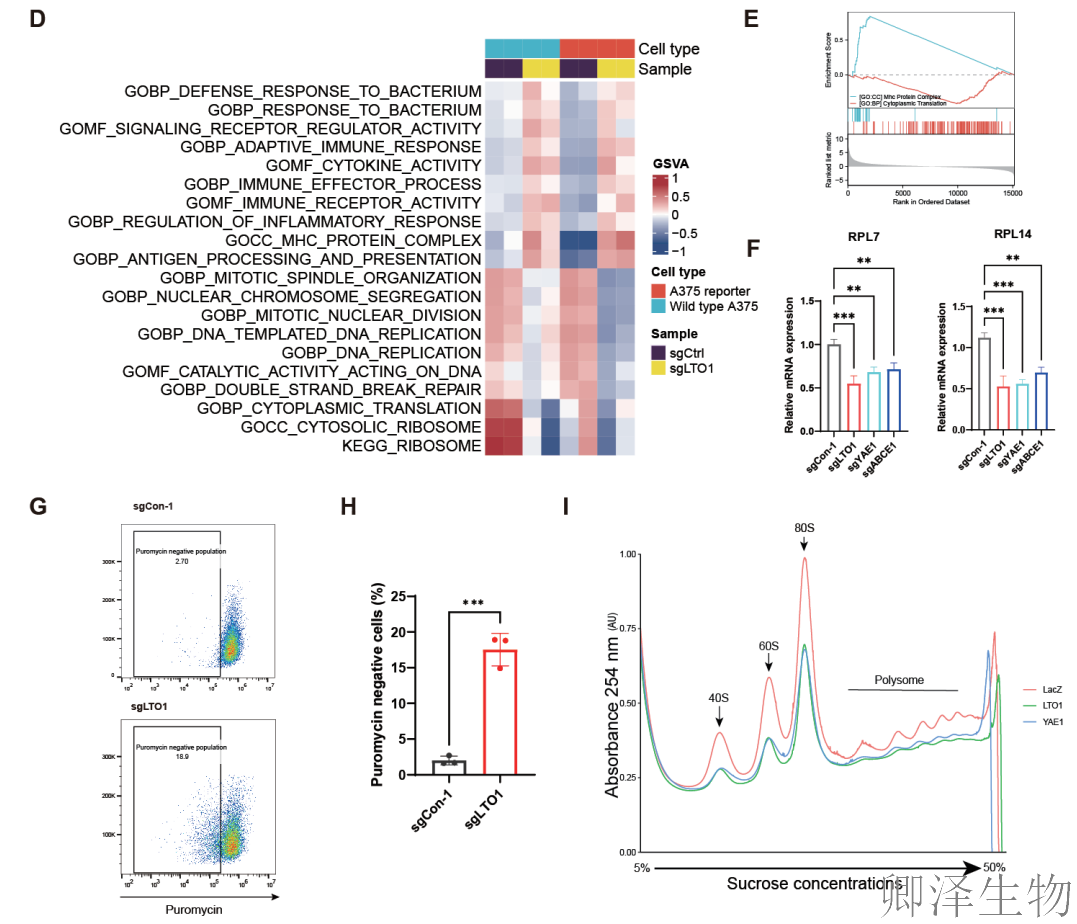
嘌呤霉素掺入实验证实(图2G-H),LTO1缺失的细胞全局蛋白质合成效率明显降低,有更多细胞处于“停产”或“低产”状态。
Polysome profiling分析提供了更清晰的图像(图2I):在缺失LTO1或YAE1的细胞中,功能完整的核糖体(80S)与未组装的“零件”(60S亚基)比例异常增高,同时高效翻译的“流水线”(多核糖体)数量减少。这双重证据表明,LTO1/YAE1的缺失,既阻碍了核糖体“零件”(60S亚基)的正确组装,也损害了整体“生产线”(翻译)的效率。
LTO1/YAE1/ABCE1缺陷增强MHC-I总体表达
转录组分析显示,敲除 LTO1、YAE1或ABCE1后,一系列与“抗原呈递”相关的基因被显著激活(图3A-D)。其中,最为关键的是MHC-I分子的编码基因(HLA-A, B, C)。流式细胞术和分子检测均证实,在多种肿瘤细胞中敲除这些基因,能实质性地上调细胞表面的MHC-I蛋白和mRNA水平。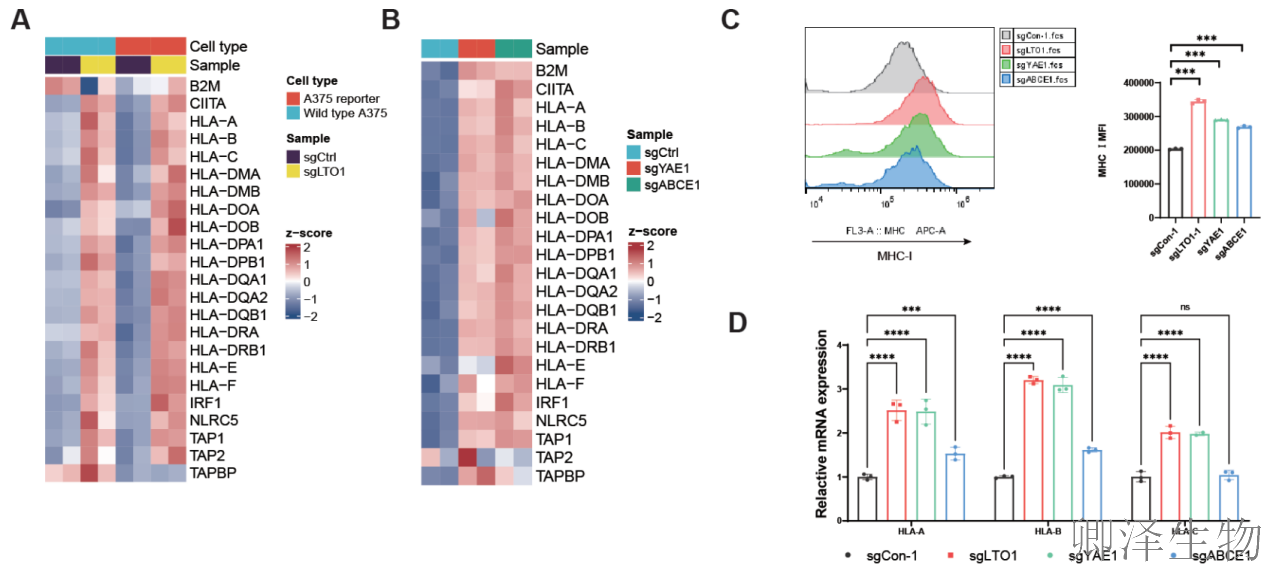
机制探索显示,多个已知能启动MHC-I表达的“总开关”转录因子(如 IRF1, NF-κB, NLRC5)及其下游的执行元件(如TAP1)在敲除细胞中均被同步激活。更有趣的是,当用免疫信号分子干扰素-γ刺激细胞时,LTO1的缺失能进一步增强MHC-I的上调效果(图3E-G)。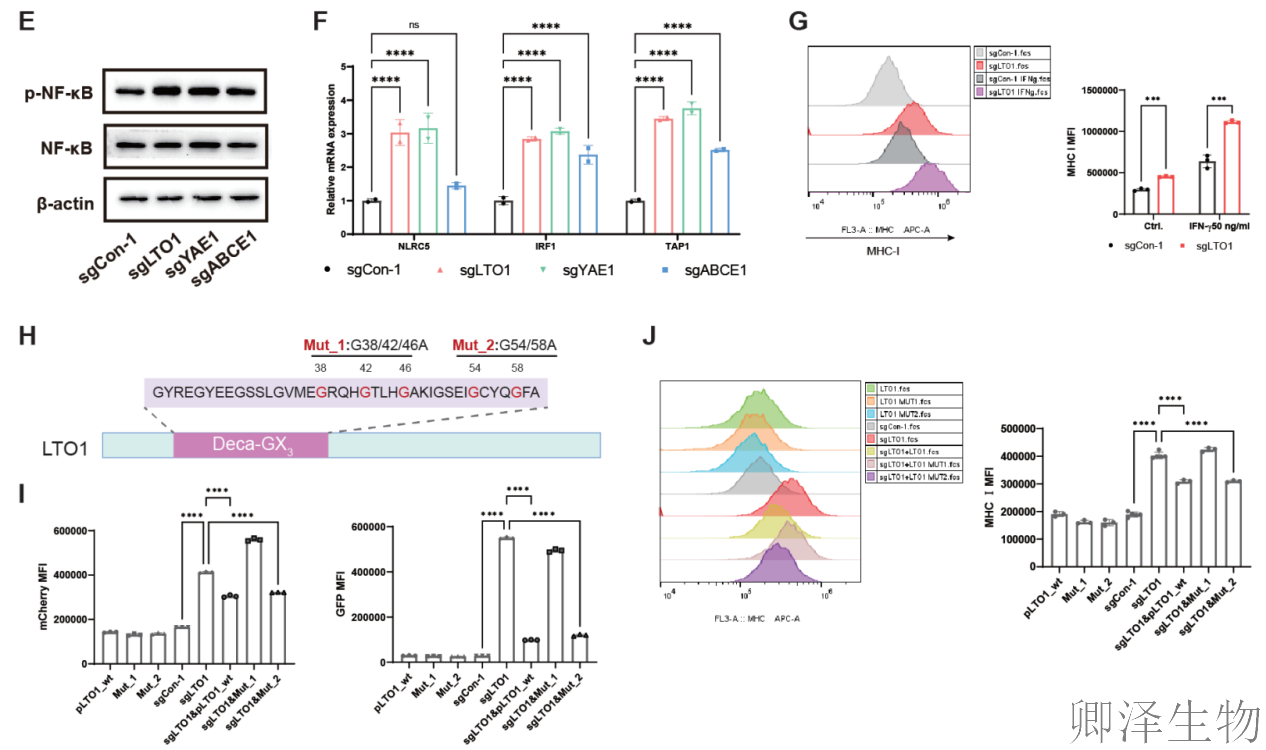
同时,研究进行了“功能回补”实验,确认上述效应是特异性地由LTO1蛋白的特定功能所介导,而非脱靶效应(图3H-J)。
LTO1和YAE1失调并与人类癌症中的免疫浸润呈负相关
随后,研究将细胞内的分子机制与宏观的临床数据联系起来。
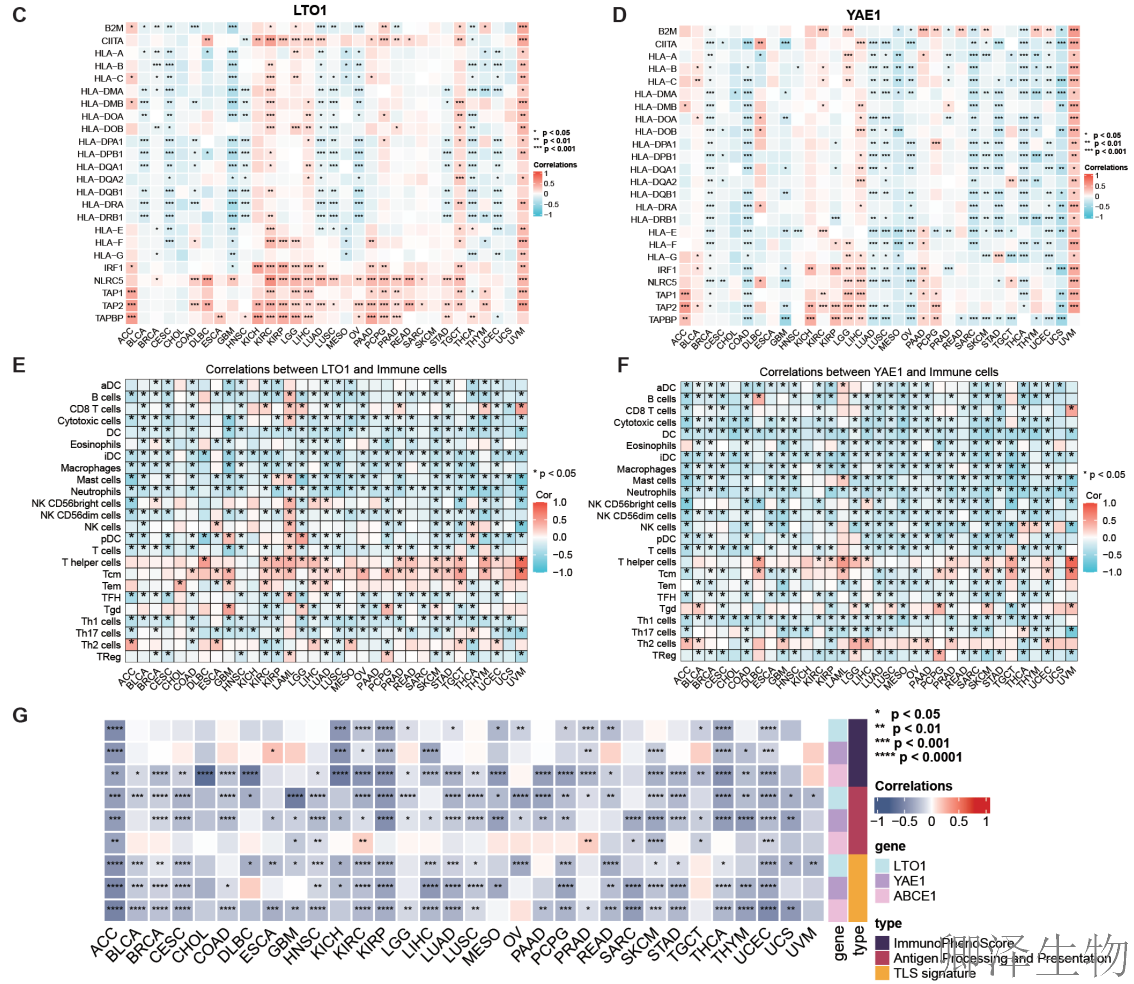
泛癌分析表明,LTO1和YAE1是潜在的“促癌”和“免疫抑制”因子。它们在多种肿瘤中高表达,且与更弱的抗肿瘤免疫反应(更低的MHC-I、更少的免疫细胞浸润、更差的免疫微环境)显著相关。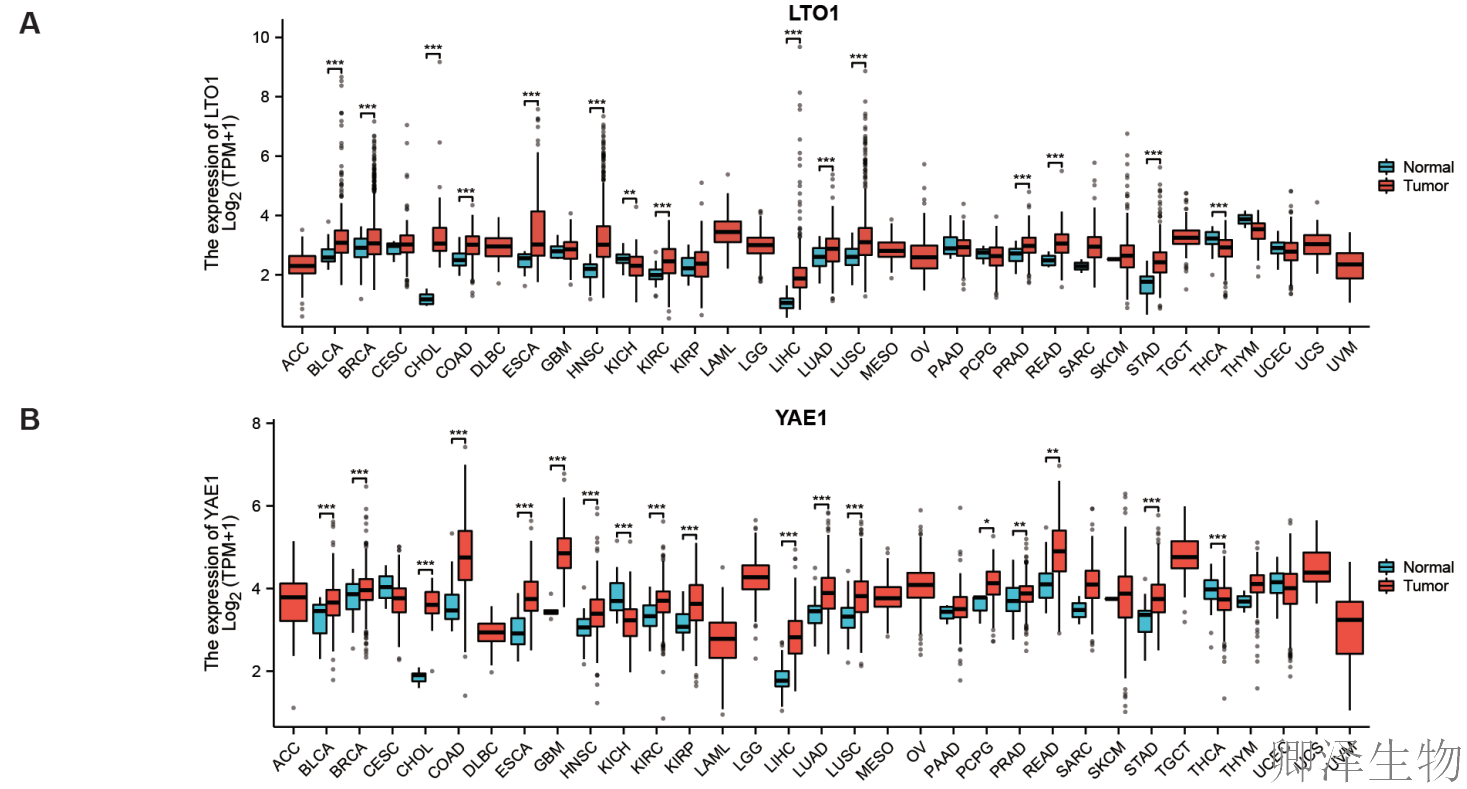
LTO1/YAE1缺陷致敏T细胞激活
前文中,看到敲除LTO1/YAE1能上调肿瘤细胞的“身份标签”MHC-I。但最关键的问题是:这真的能让免疫系统(特别是T细胞)更好地识别和攻击肿瘤吗?
研究构建了一个巧妙的“T细胞激活探测器”。这些细胞被改装了能识别特定肿瘤抗原(NY-ESO-1)的T细胞受体(1G4 TCR),并且一旦T细胞受体被激活,就会启动一个绿色荧光蛋白(ZsGreen)的表达。这样,T细胞是否“看到”肿瘤细胞,就会直接显示为“发绿光”(图5A-C)。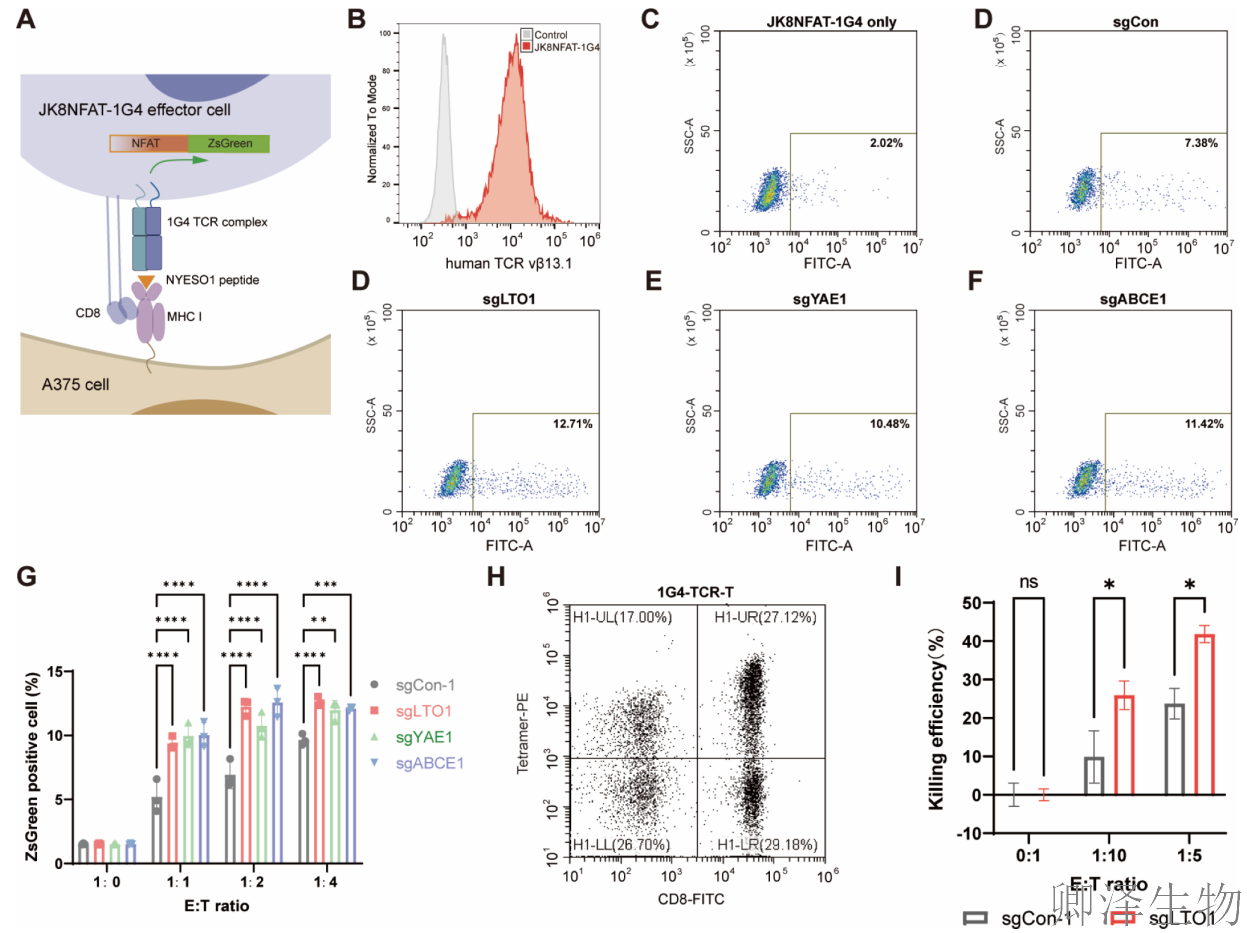
这个“探测器”去识别经过处理的A375肿瘤细胞,发现:与普通的A375细胞共培养时,一部分T细胞被激活(发绿光)。但当与敲除了LTO1、YAE1或ABCE1的A375细胞共培养时,被激活的T细胞比例显著增加(图5D-G)。这直接证明,降低LTO1/YAE1的表达,能显著增强T细胞对肿瘤细胞的识别和响应。
另外,在“TCR-T细胞疗法”的体外模型中,从人血液中提取免疫细胞,并为其装备了同样的NY-ESO-1特异性TCR,制成了“精准杀伤部队”。杀伤实验显示,在多个不同的“兵力”(T细胞)与“目标”(肿瘤细胞)比例下,LTO1缺失的肿瘤细胞都被TCR-T细胞杀死的效率更高(图5H-I)。
综上,通过基因敲除等手段抑制LTO1/YAE1,不仅能从分子和细胞水平上调MHC-I,更能实实在在地增强T细胞对肿瘤的识别和杀伤能力。
受损的NMD诱导MHC-I上调并增强免疫细胞激活
MHC-I上调,是LTO1/YAE1这个“零件”坏掉导致的特例,还是整个“质检系统”(NMD)出问题的普遍结果?
已知NMD核心因子SMG6和SMG7的敲除,以及NMD抑制剂处理,均能显著升高MHC-I表达(图6A-D)。证实了NMD与MHC-I之间的因果关系。在测试的药物中,铁螯合剂抑制NMD和上调MHC-I的效果存在明显的剂量依赖性,且具有广谱性(图6E-I)。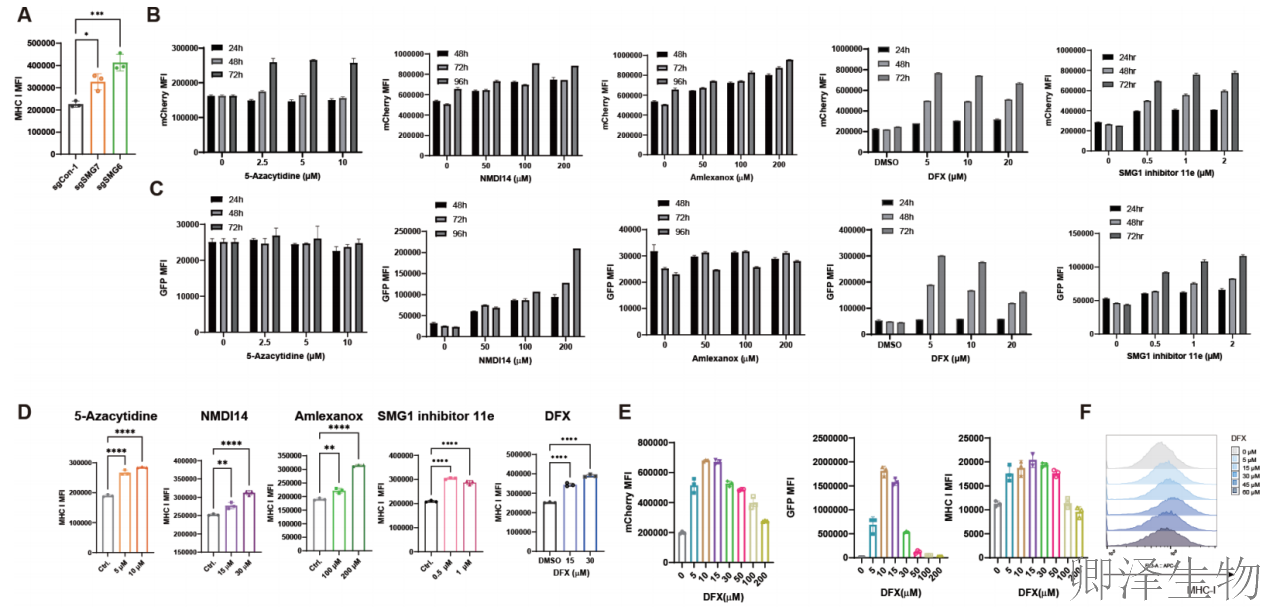
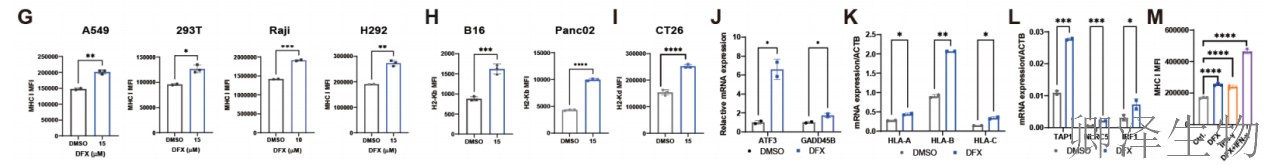
分子层面,铁螯合剂处理同样会导致已知NMD靶标mRNA积累,并激活NLRC5、IRF1等MHC-I上游调控因子。与免疫信号分子干扰素-γ联用,能进一步协同提升MHC-I水平(图6J-M)。
至此,系列实验完成了从“基因特例”到“通路共性”的论证,并成功找到了可模拟该表型的工具药物。
低剂量铁螯合剂增强免疫细胞识别和激活
铁螯合剂,真的能让免疫系统更有效地识别和攻击肿瘤吗?研究通过从体外到体内、从细胞实验到动物模型的完整证据链:
体外验证——增强T细胞“开关”(图7A-C)
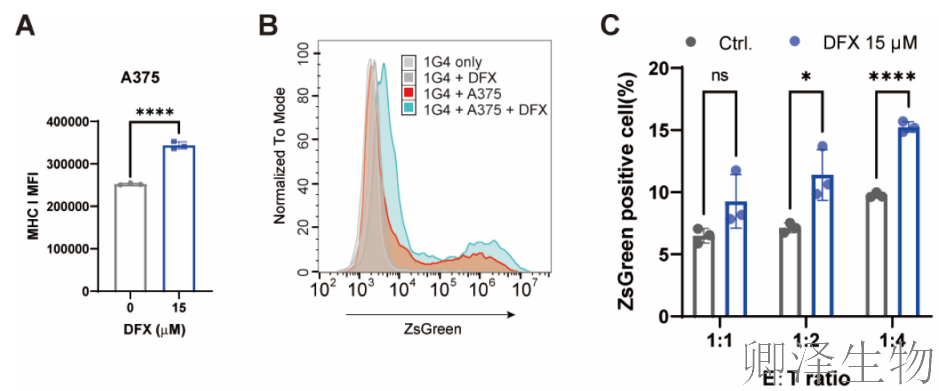
小鼠模型验证——提升T细胞“杀伤力”(图7D-E)
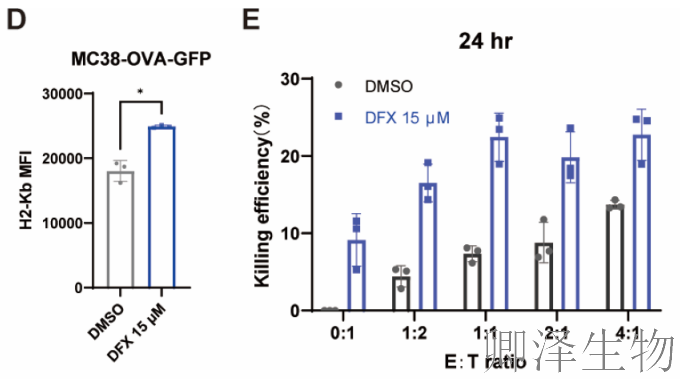
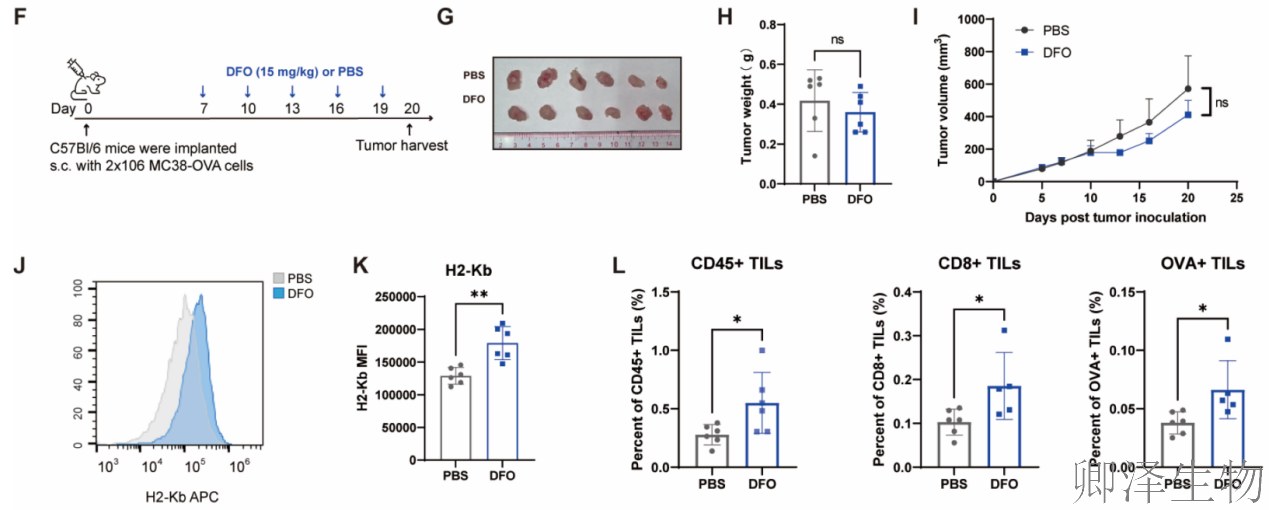
体内效果——改变肿瘤“微环境”(图7F-L)
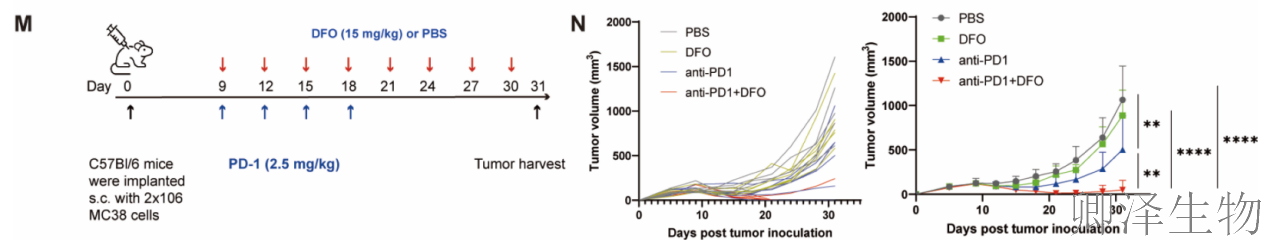
联合治疗——“破锁”与“松刹车”双管齐下(图7M-N)
结论
这项研究首次确立了LTO1/YAE1复合物通过NMD通路负调控MHC-I表达的新功能,揭示了肿瘤细胞利用基础mRNA监控机制来逃避免疫监视的一条全新途径。更重要的是,它提出了一个极具转化潜力的治疗策略:靶向NMD(例如使用铁螯合剂)可以作为一种“免疫增敏”手段,与现有的T细胞疗法(如TCR-T)或免疫检查点阻断疗法联合使用,有望逆转肿瘤的免疫“冷”状态,为对现有免疫疗法不响应的患者提供新的治疗选择。









