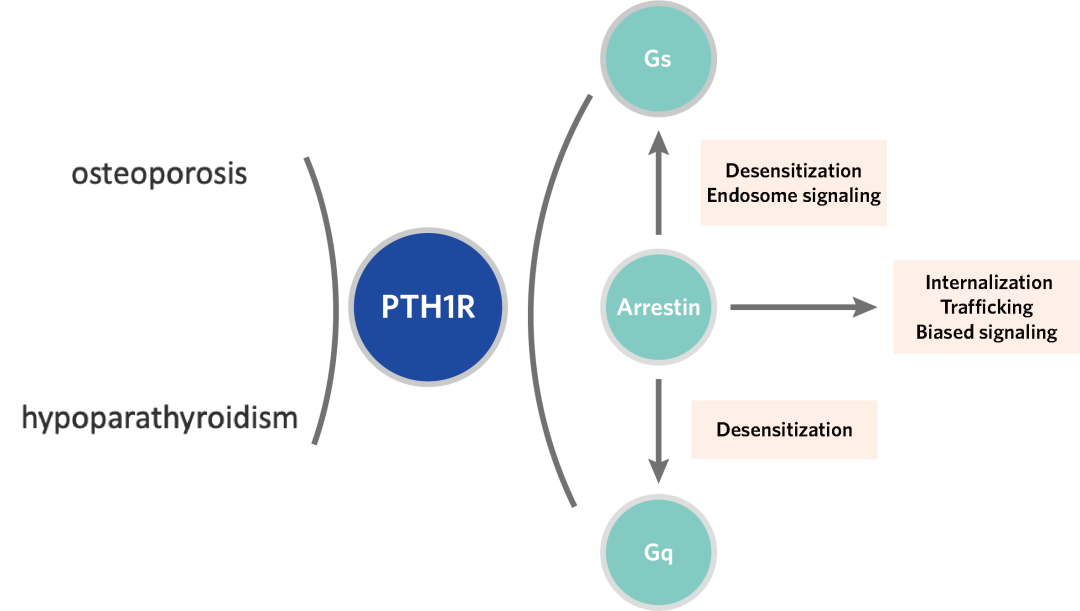
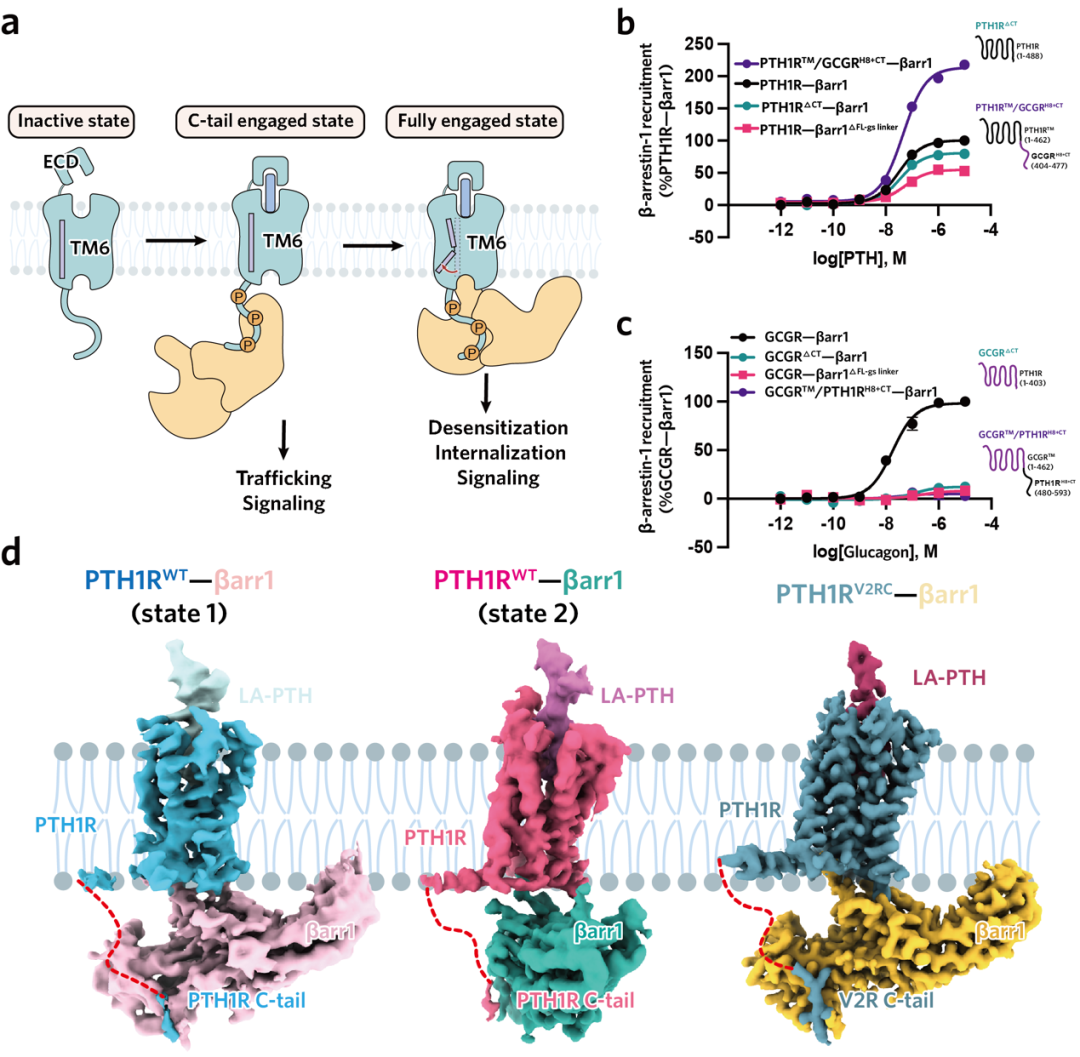
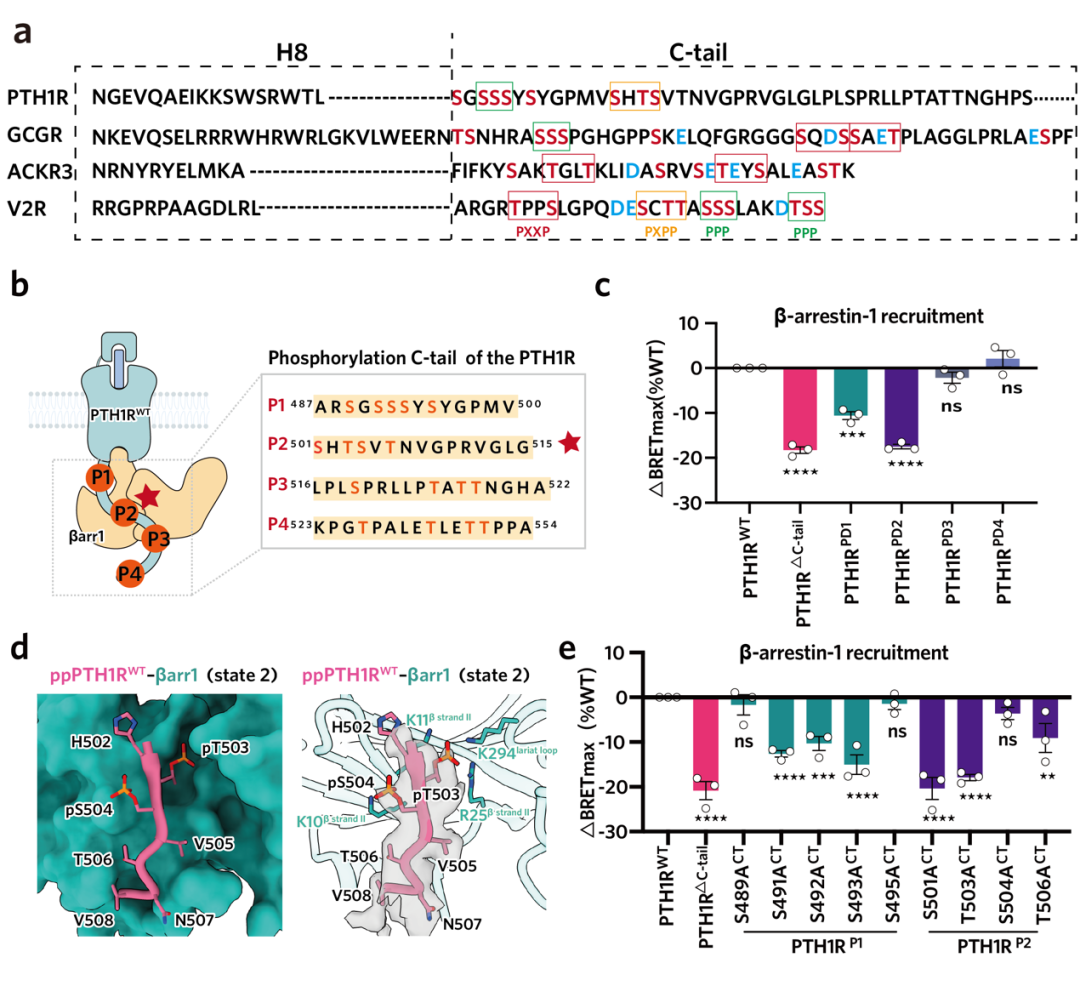
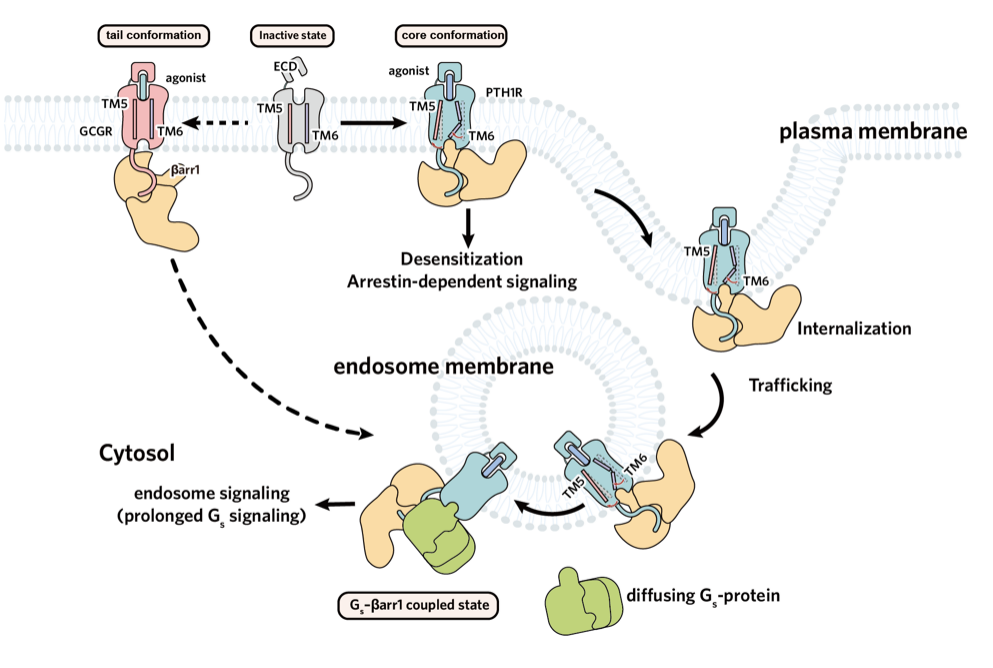
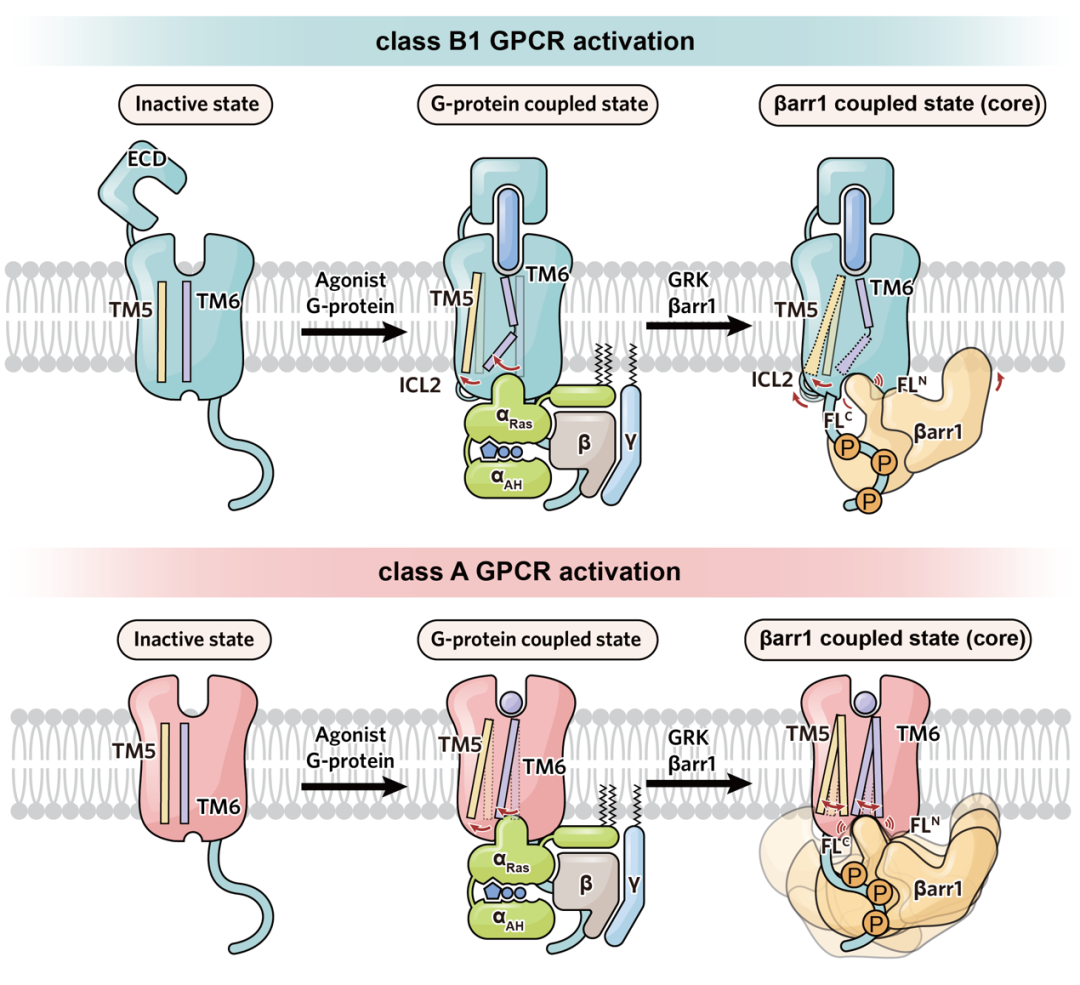
近年来,慢性肾脏病(CKD)发病率持续攀升,已成为不容忽视的公共卫生问题。其严重并发症 — 慢性肾脏病矿物质与骨异常(CKD-MBD),是导致 CKD 患者致残、致死等不良结局的重要原因之一。目前,针对 CKD-MBD 的临床用药普遍缺乏对信号通路的精准选择性,导致促成骨作用受限且副作用明显。因此,为破解当前的临床困境,亟待开发能精准靶向有益信号通路的创新药物。 为此,东部战区总医院国家肾脏疾病临床医学研究中心刘志红院士团队与浙江大学良渚实验室张岩教授团队开展紧密合作,聚焦于维持钙磷平衡及促进骨骼合成的药物靶点 —— 甲状旁腺激素 1 型受体(PTH1R)。通过深度解析其配体结合与下游信号转导的分子机制,助力新型靶向药物的理性设计,为肾性骨病的治疗开辟全新路径。 目前以 PTH1R 为靶点的上市药物,虽能有效促进骨合成,却常因引发高血钙等副作用而应用受限。PTH1R 疗效与副作用的产生来源于其独特的配体结合及下游信号转导机制。PTH1R 可同时介导短时程与长时程的 cAMP 信号升高,而后者正是导致高血钙等不良反应的关键。现有研究认为,长效配体的持续结合以及阻遏蛋白(β-arrestin)介导受体内吞至内体后所产生的「内体信号」,共同维持了这种长时程的 cAMP 信号(图 1)。为阐明信号时程差异的分子机制,研究团队通过解析长效与短效配体-PTH1R-Gs 复合物的高分辨率结构结合功能试验验证,揭示了控制信号时长的关键氨基酸位点,并据此设计出时程可控的新分子,相关成果于 2022 年发表在《Nature Communications》1。 然而,仅基于现有受体结合口袋的改造,仍不足以实现临床「增效减毒」的根本性突破。真正的核心瓶颈在于下游信号调控:由于 β-arrestin 与 GPCR 的结合具有高度灵活性,获取高分辨率的 PTH1R–β-arrestin 复合物结构一直是领域内亟待解决的难题。这一关键结构的缺失,严重阻碍了基于完整信号通路的药物理性设计。 ▲图 1. PTH1R 作用机制及药物作用靶点 2026 年 4 月 25 日,东部战区总医院国家肾脏疾病临床医学研究中心刘志红院士团队与浙江大学良渚实验室张岩教授团队开展跨学科联合攻关,在《Nature Communications》上发表了题为《Core Conformation of Arrestin Coupling to Parathyroid Hormone Type 1 Receptor》2 的重要研究成果(图 2)。该研究首次成功解析了 PTH1R 与 β-arrestin1(βarr1)的激活态复合物三维结构,在原子层面揭示了这一关键相互作用模式。这一突破性结构实现了从分子解析到临床药理的跨维度链接,为完整阐释 PTH1R 从配体结合到偶联下游信号蛋白的全流程提供了决定性框架,使得在分子层面精确解析 β-arrestin 调控的 PTH1R 内体长效信号转导及其介导的偏向性信号转导成为可能,从而为开发新一代靶向药物奠定了坚实的结构基础。 ▲图 2. Nature Communications 文章封面 本项研究突破 1、首次揭示了野生型 PTH1R 与 βarr1 偶联的两种不同的核心构象,为理解 β-arrestin 与 B1 类 GPCR 的相互作用模式提供了全新视角 在 GPCR 的信号转导研究中,class A GPCR 与 β-arrestin 的相互作用虽呈多样化,但均受控于经典的核心结合构象。相比之下,class B1 GPCR 虽具备高度的家族保守性及高度相似的 G 蛋白偶联模式,但其与 β-arrestin 的结合机制长期成谜。迄今为止,该家族仅解析了 GCGR 与 β-arrestin 的复合物结构,且显示出异于经典的「尾部结合」模式。这种依赖 H8 螺旋和 C 末端的特殊构象是否为 class B1 GPCR 所共有?基于这一关键科学问题,我们前期的功能学验证明确提示,PTH1R 具有与 GCGR 迥异的 β-arrestin 结合特性。针对该现象,团队突破了复合物制备的技术壁垒,历经大量蛋白纯化方案筛选与数据计算策略优化,最终成功捕捉并解析了两种处于核心构象结合模式的 PTH1R–β-arrestin 复合物结构(图 3)。结构解析证实,PTH1R 与 βarr1 的偶联高度依赖核心区域的相互作用。这一发现揭示了同家族受体在 β-arrestin 招募策略上存在显著的机制分化。更为重要的是,在所解析的核心构象中,β-arrestin 呈现出两种截然不同的装配模式,深入确证了其在靶向识别受体时的构象可塑性与结合模式的多样性。这些精细的差异化机制不仅填补了领域内的结构空白,更直接为研发靶向特定信号通路的「偏向性药物(Biased ligands)」提供了坚实的结构基石,从而有望从底层逻辑上克服传统药物的副作用,实现精准、安全的临床干预。 ▲图 3. PTH1R–βarr1 高分辨率结构 2、阐明了 PTH1R C 端尾部磷酸化编码 GPCR C 末端尾部(C-tail)的特异性「 磷酸化密码」是调控 β-arrestin 招募模式及下游生理功能的关键决定因素。研究表明,PTH1R 的 C-tail 独特的磷酸化编码与 β-arrestin 紧密结合,因此可以介导其独特的内体信号:通过 β-arrestin 与受体 C 末端尾部的特异性结合,受体跨膜核心得以释放并持续偶联 Gs 蛋白,从而介导从细胞膜至内体膜的长时程 cAMP 信号。为阐明这一独特信号模式的底层逻辑,精确解析其磷酸化编码并明确其尾部构象转换机制显得尤为迫切。针对这一关键问题,本研究结合冷冻电镜结构解析与定点突变功能学验证,首次揭示了 PTH1R 的 C 末端尾部是通过高度保守的「P-X-P-P」磷酸化基序介导 β-arrestin 的招募与效应发挥(图 4)。该基序的结构确证,不仅填补了 PTH1R 磷酸化识别机制的空白,也为后续模拟不同磷酸化状态、设计具有偏向性信号调控功能的 PTH1R 创新分子提供了关键的结构蓝图。 ▲图 4. PTH1R 的 C 端尾部磷酸化编码 3、明确了 β-arrestin 核心构象在介导 PTH1R 内吞和转运以促进持续内体信号中的关键作用,关联了结构发现与信号转导 βarr1 可以通过核心构象和尾部构象两种方式与 GPCR 结合,进而参与介导受体信号转导及生理功能过程。既往研究认为 βarr1 的核心构象更多的参与受体的 G 蛋白脱敏及偏向性信号转导,而尾部构象则参与了受体的内吞和运输至内体。而我们通过结构分析确定 βarr1 和 PTH1R 核心相互作用的关键氨基酸,通过检测其内吞、运输功能,证实 βarr1 核心构象调控 PTH1R 的内化、运输和内体信号(图 5)。进一步的结构分析表明,在核心结合构象中,βarr1 的 C 边缘脱离受体并与膜相互作用,其暴露的 C 边缘更易于与网格蛋白和 AP2 结合,从而促进形成稳定的 PTH1R–βarr1– 网格蛋白 –AP2 复合物驱动受体内吞,这些发现为 βarr1 核心构象在调控内体信号转导中的基本生理作用提供了有力证据。 ▲图 5. PTH1R 激活结合下游信号蛋白全过程 4、揭示了 β-arrestin 结合 GPCRs 核心构象的保守性与多样性 我们解析的 βarr1 与 PTH1R 复合物结构揭示了 βarr1 与 B1 类和 A 类 GPCRs 在核心构象偶联模式上的重要差异。研究发现,A 类 GPCRs 的转导蛋白结合口袋具有高度可变性,这使得阻遏蛋白能够以多种异质性模式结合。相比之下,我们的结果表明 B1 类 GPCRs 在阻遏蛋白的核心结合中表现出显著的结构相似性,其典型特征包括 TM6 的显著弯曲和一个高度保守的结合口袋(图 6)。这项研究深化了我们对不同转导蛋白如何与 GPCRs 偶联以及如何调控下游信号转导机制的理解。 ▲图 6. β-arrestin 结合 class A GPCR 多样性和 class B GPCR 保守性 研究意义 本研究不仅首次揭示了 PTH1R 与 β-arrestin 结合的独特构象,为其偏向性信号转导机制研究提供了原子级蓝图,更为代谢性骨病的精准治疗提供了结构生物学新范式与药物设计新策略。研究证明了「配体-Gs 复合物解析」与「受体-β-arrestin 复合物解析」相结合的全流程结构解析策略,是阐明 GPCR 复杂时空调控网络、破解「信号双时相性」分子基础的关键路径。这为未来开发 G 蛋白偏向性或 β-arrestin 偏向性的配体提供了直接的结构依据,使得 「在保留促骨合成疗效的同时,精准规避副作用通路」这一临床理想成为可能,从而为 CKD-MBD、骨质疏松等疾病的个体化药物研发与精准干预提供了全新的科学视角和坚实的理论支撑。 作者介绍 东部战区总医院国家肾脏疾病临床医学研究中心刘志红院士和浙江大学良渚实验室张岩教授、毛春友研究员为论文的共同通讯作者,东部战区总医院国家肾脏疾病临床医学研究中心翟修文博士、王钢博士,浙江大学良渚实验室郭嘉博士、沈庆亚博士和陈礼楠博士为共同第一作者。 全文链接: 1. Molecular insights into the distinct signaling duration for the peptide-induced PTH1R activation「https://www.nature.com/articles/s41467-022-34009-x」 2. Core Conformation of Arrestin Coupling to Parathyroid Hormone Type 1 Receptor「https://www.nature.com/articles/s41467-026-72448-y」