

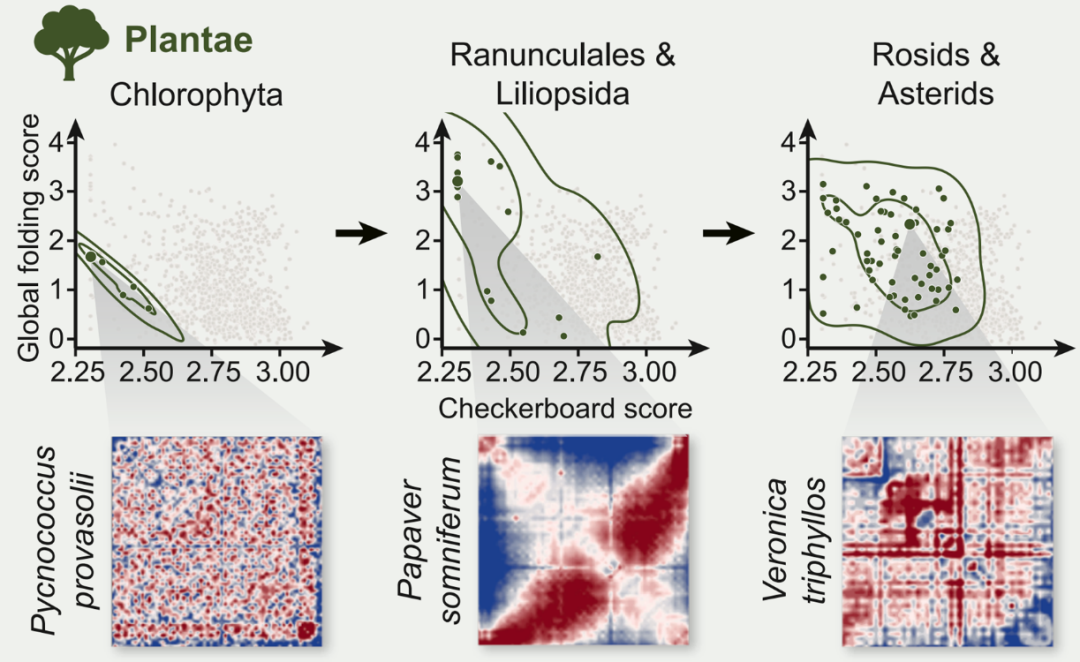
1953 年,沃森和克里克揭示的 DNA 双螺旋结构是生命演化史上最重要的发现之一。这一发现奠定了现代分子生物学的基石,也让我们理解了「结构决定功能」这一核心原理。然而 70 多年过去,当科学家们将目光从单一的 DNA 分子转向整个基因组的三维空间组织时,一个根本性的问题仍未得到解答:基因组在细胞核内的空间折叠,是否也遵循着某种普适的演化规律? 2026 年 4 月 21 日,西安交通大学叶凯团队在 Cell 在线发表题为「The evolution of high-order genome architecture revealed from 1,000 species」的研究论文,该研究不仅首次描绘了跨越 38 亿年生命史的高维基因组进化图谱,更发现了一个出人意料的事实 —— 植物和动物在基因组空间组织上选择了截然不同的策略,却殊途同归地服务于同一个生物学目的。 从研究方法来看,这是一项在规模上前所未有的尝试。研究团队整合了来自达尔文生命树项目的 Hi-C 数据,涵盖了 1,025 个物种,时间跨度约 38 亿年,涉及从基因组仅约 2.1 Mb 的古菌到基因组规模高达约 14.6 Gb 的六倍体小麦,从单细胞细菌到复杂的多细胞哺乳动物等不同层级的生命形式。面对基因组大小差异如此巨大的数据集,传统分析方法完全失效。为此,研究团队专门开发了 NormDis 算法,通过距离依赖性偏差校正和缩放采样策略,首次实现了跨物种的三维基因组结构比较。 千种物种、38 亿年演化:两种高阶结构主宰基因组折叠 分析结果揭示出两种主导的高阶基因组结构。第一种被称为「全局折叠」,描述的是整套染色体在细胞核内的宏观空间排布,通常由着丝粒和端粒作为锚点将染色体锚定在特定位置。第二种是「棋盘格」结构,表现为 Hi-C 接触图谱上规则的交叉纹理,反映的是常染色质和异染色质区域的空间区室化。有趣的是,这两种结构在生物类群中的分布呈现出鲜明的分化趋势。 植物选择了强化全局折叠的策略。数据显示,无论是单子叶植物还是双子叶植物,都显著偏好一种称为「中心-末端轴」的折叠模式 —— 着丝粒和端粒分别聚集在细胞核的不同区域,染色体臂则平行排列。与此同时,植物的棋盘格结构相对较弱。这种结构选择与基因在染色体上的线性排列密切相关:研究发现,植物倾向于将属于同一代谢通路的基因成簇排列在同一条染色体上,并通过全局折叠将这些线性相邻的基因在空间上拉近,从而实现协同调控。 动物则走上了另一条道路。随着生物复杂性的增加,动物逐步发展出越来越强的棋盘格结构。从最原始的多细胞动物海绵开始,基因组已呈现出初步的棋盘格结构;至哺乳动物阶段,这种区室化在分析类群中达到最强。相比之下,动物的全局折叠较弱,基因在染色体上的线性聚类也不明显。这意味着动物采用了不同的调控策略 —— 不依赖基因在 DNA 序列上的相邻,而是通过精细的空间区室化将功能相关的调控元件和靶基因组织到物理邻近的空间环境中。 殊途同归:植物靠「线性聚类」,动物靠「空间区室化」 更耐人寻味的是人类早期胚胎发育中的动态转换。在二细胞期这一全能性状态下,胚胎基因组呈现出类似早期生命形式的特征 —— 强全局折叠、弱 棋盘格。随着发育推进到囊胚期,结构发生戏剧性转换:全局折叠减弱,棋盘格显著增强。这一转换恰好与细胞从全能性向谱系特化的转变同步,提示基因组结构的重组可能与细胞命运决定相关。 胚胎发育的缩影:二细胞期到囊胚期,重演 38 亿年的结构演化 这项研究的重要意义在于,它将「结构决定功能」这一原理从 DNA 双螺旋层面拓展到了全基因组的三维空间组织层面。植物和动物独立进化出多细胞性,却选择了不同的基因组折叠策略:植物投资全局折叠和线性基因聚类,换取调控效率;动物投资精细的区室化,换取调控灵活性。两种策略看似对立,实则都服务于同一个核心生物学功能 —— 通过空间组织实现基因表达的精确调控。 这一发现也为理解生命多样性提供了新的维度。为何植物能产生如此丰富的次生代谢物?或许与其基因线性聚类的结构优势有关。为何动物能演化出如此复杂的神经系统?或许与其精细的区室化调控能力有关。未来,通过人为调控基因组的空间结构,或许能在作物改良中优化代谢通路效率,或在再生医学中提高细胞重编程的成功率。 不过,这项研究仍存在局限。由于测序深度和基因组大小的差异,研究团队无法在所有物种中解析出精细的三维结构,如拓扑关联结构域和染色质环。大部分数据来自单一组织类型,且缺乏单细胞分辨率。此外,这些高阶结构形成的分子机制,特别是染色体结构维持蛋白和染色质修饰的具体作用,仍需更多功能实验来阐明。 原文链接: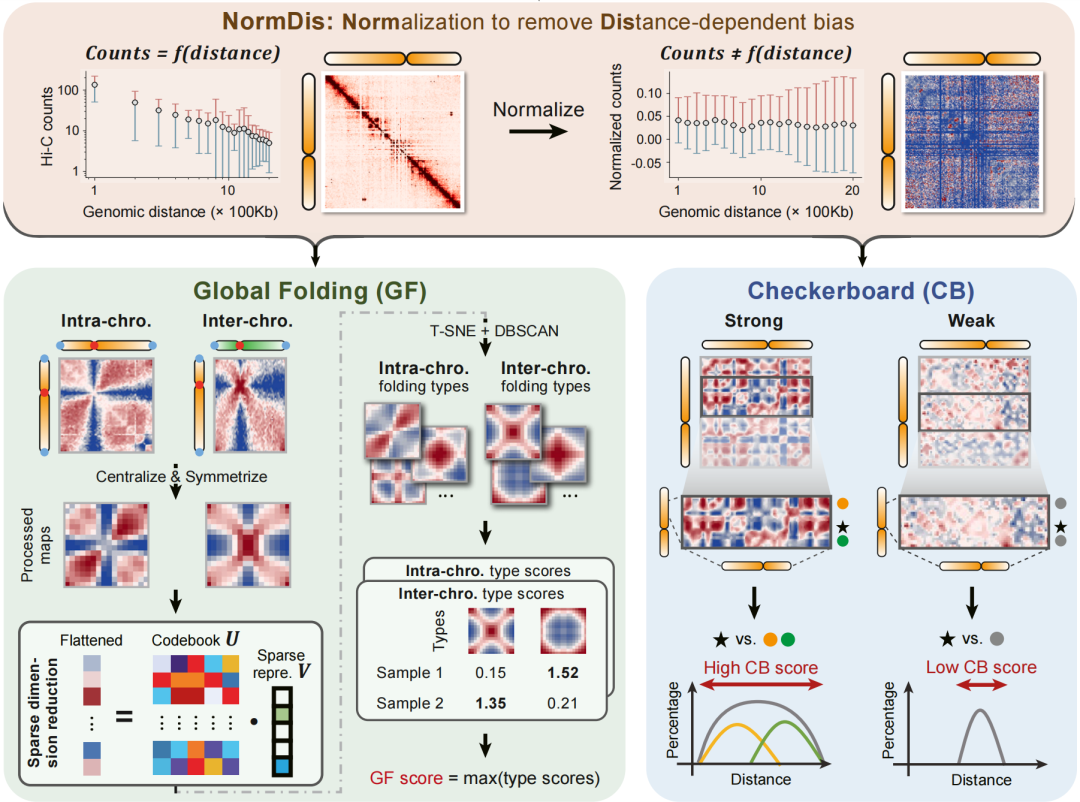
https://doi.org/10.1016/j.cell.2026.03.042









